situations, calcium acts both as a signal transmitter from the outside of the cell to the inside and as an activator or stabilizer of the functional proteins involved. In fact, ionized calcium is the most common signal transmitter in all of biology. It operates from bacterial cells all the way up to cells of highly specialized tissues in higher mammals.
calcium. Part is excreted in the urine through the kidney, and part enters the slower exchange pools of soft tissue and bone. Dietary calcium influences calcium absorption and, consequently, fecal calcium and, to a lesser extent, urinary calcium excretion. An obligatory loss of calcium occurs through endogenous secretion, urine, and skin. Gender, age, and racial differences in calcium metabolism exist. Adolescents are more efficient at using calcium than are young adults, and elderly persons are the least efficient. Boys are more efficient at calcium metabolism than girls, and blacks are more efficient than whites.
TABLE 7.1 CALCIUM (Ca) METABOLISM AS INFLUENCED BY RACE AND AGE | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
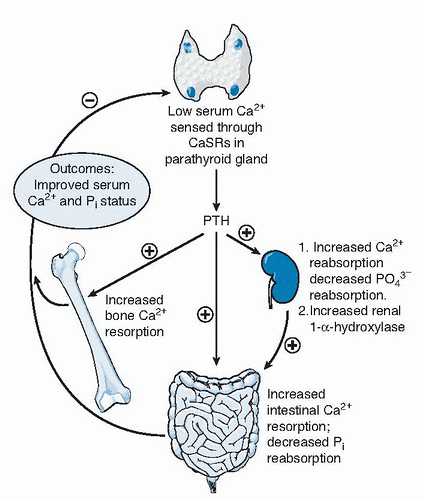 Fig. 7.1. Homeostatic regulation of calcium (Ca2+) depicting the changes in vitamin D and parathyroid hormone (PTH) when plasma calcium falls to less than 2.5 mM. CaSR, calcium sensing receptor; Pi, inorganic phosphate; PO43-, phosphate. |
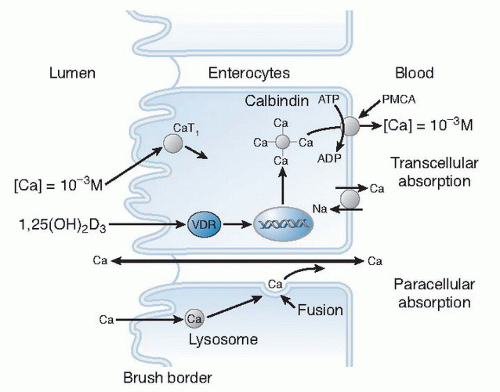 Fig. 7.2. Calcium (Ca) absorption showing active, transcellular absorption and passive, paracellular absorption. Paracellular absorption is bidirectional; transcellular absorption is unidirectional. Ca enters the cytosol down a concentration gradient. Ca enters the cell through CaT1 and is transported across the enterocyte against an uphill gradient with the aid of vitamin D-induced calbindin, probably at least partially through endosomes and lysosomes. Finally, it is extruded at the basolateral membrane primarily by the plasma membrane calcium adenosine triphosphatase (ATPase) pump (PMCA) and secondarily by the sodium (Na+)/Ca2+ exchanger or by exocytosis. ADP, adenosine diphosphate; VDR, vitamin D receptor. |
Transcellular: This saturable (active) transfer involves a calcium-binding protein, calbindin.
Paracellular: This nonsaturable (diffusional) transfer is a linear function of calcium content of the chyme.
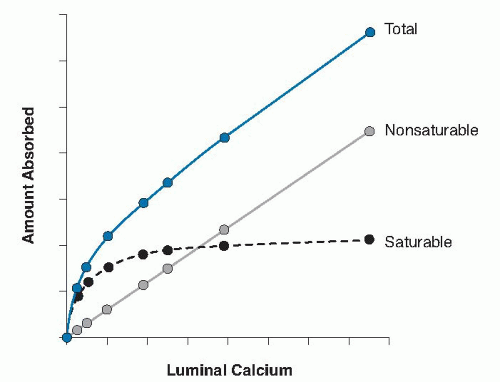 Fig. 7.3. Calcium is absorbed by both saturable and nonsaturable pathways. Total calcium transport (the sum of a saturable component [A] defined by the Michaelis-Menten equation and a concentrationdependent, nonsaturable component [B] defined by a linear equation) is described by a curvilinear function. |
in young premenopausal women (25). The age-related decrease in calcium absorption from intestinal resistance to 1,25(OH)2D3 has been associated with decreased VDR levels (26), as well as with reduced estrogen levels (23).
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


