Long before the musculoskeletal system emerged in the geological scheme of things, flexible multicellular organisms splashed about in the waters of the world. First came cartilage and then, about 1.5 billion years ago, a violent event changed evolution. Movements in the earth created huge amounts of minerals that washed into the sea. What followed? Spines, shells, and hard body parts. Limbs were extended and protective shields created. Calcium carbonate was replaced by calcium phosphate and then calcium hydroxyapatite, a building block of great chemical stability resistant to acidic environments. The great evolutionary leap was underway (1).
Bone consists of apatite crystals embedded within collagen and bathed in bone-promoting proteins. It is initially formed by a combination of endochondral ossification and membranous ossification (MO). Continuously shaped and reshaped throughout life (“modeling”), its basic unit of turnover is a microscopic coupled event linking osteoclasts and osteoblasts (“remodeling”). Both measurable influences such as hormonal activity of parathormone (parathyroid hormone, PTH) and less measurable events such as electrostatic forces are critical. Local cellular events and tissue factors, including cytokines, chemokines, matrix metalloproteinases (MMPs), ligands, and signaling transduction cascades, are important regulators.
The constituents of bone are a composite of soft flexible collagen fibrils and hard inflexible calcium and phosphorus crystals, a composition that is stronger and lighter than either constituency alone. Embedded in this matrix are bone-forming proteins, bone-related proteins, and both blood and mesenchymally derived cells, which differentiate into osteoclasts and osteoblasts, respectively. In addition to the bone tissue proper, ancillary soft tissues such as fatty and hematopoietic marrow; surface fibrous tissue called periosteum; and associated tendons, ligaments, synovium, and joint capsules create a complex organ of ongoing mechanical, bioelectrical, and physiochemical activity.
Anthropologic studies best delineate the dynamic and dramatic modeling changes that can occur in skeletal shape, size, and density. Spear-throwing hunters from an earlier age were characterized morphologically by a specific enlargement of the ulna, not unlike that found in a current young baseball pitcher. A life characterized by squatting is evidenced by abnormal grooves in the lower end of the tibia and at the hip and knee, findings common in the skeletons of the Maoris of New Zealand and Australian aborigines. Other specific indications of skeletal changes are the degenerative joint disease of the spine in Acapulco cliff divers, abnormalities of the lower jaw attributed to clarinet playing, golfer’s toe, and dog walker’s elbow. In fact, more than 140 established skeletal markers of occupational stress have been defined (2). Forensic anthropologists have utilized skeletal changes in identifying cultural habits even in ancient times. The enlargement of finger bones in 3,000-year-old Egyptian mummies suggests flexion activity of ancient scribes.
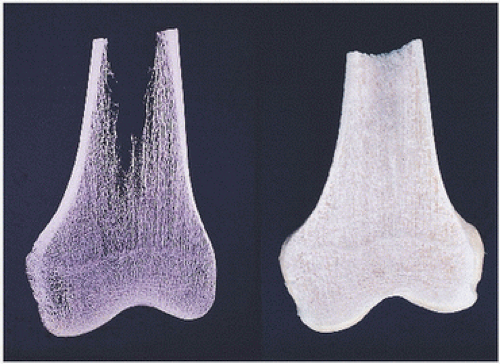
Bone is known to respond to stresses and strains by intensive modeling (macroscopic reorganization of skeletal structure) and remodeling (microscopic remodeling activity of osteoblast formation and osteoclast resorption). These are most easily seen in changes of cancellous bone macroarchitecture, as evidenced by x-rays and the gross appearance of macerated skeletal specimens (Fig. 1.1).
In a normal-sized human being, the skeleton accounts for about 15 percent of the total weight, and if water and fat are removed, the dry skeletal weight composes roughly less than 10 percent. Of this, approximately 65 to 70 percent is constituted by the mineral component of bone, and 30 to 35 percent dry organic matter. Water accounts for 5 to 10 percent of bone, and lipids less than 3 percent. Much of the latter is constituted by collagen and noncollagenized bone matrix, proteins such as bone sialoprotein, osteonectin, osteocalcin, proteoglycan, and other tissue components. The mineral component is primarily consistent of phosphorus and calcium (18 and 38 percent) and is remarkably constant. Whether one is dealing with porous or compact (i.e., trabecular or cortical) bone, the ratio of calcium to phosphorus is approximately 2:1, ratios characteristic of hydroxyapatite crystals, the main crystalline component of bone.
FIGURE 1.1. Bone structure, distal femur. Bone consists of a cortical bone envelope within which are interconnecting horizontal and vertical systems of cancellous (trabecular) bone. Macerated gross specimen of bone (right). Specimen roentgenogram (left).
Collagen
Collagen is the major extracellular matrix of bone and acts as the scaffold for crystallization of calcium hydroxyapatite crystals, causing the unique flexible and inflexible properties of bone tissue. Its ubiquitous presence in connective tissue is manifested by a broad range of biomechanical and other functions of skeletal tissue. In bone, it provides the scaffold on which mineral is deposited, counterbalancing the brittleness of the mineral by its tensile strength.
Tendons, almost exclusively made up of collagen, serve a different function, as does fibrocartilage, which has a tensile strength of collagen fibrils. Tendons provide a compressive resistant tissue in opposition to the tendency of the highly charged proteoglycans of articular cartilage to generate a swelling type of pressure.
The broad functions of collagen are manifested in the different types of collagen and indeed the ability of modifications in collagen type to occur from tissue to tissue, and, in some instances, within the same tissue after a reaction such as fracture.
Collagen is a triple helical fibrillary protein structure (3) (Fig. 1.2). Its three-dimensional polypeptide orientation may be altered by the presence of certain amino acids, giving some types of collagen particular characteristics, such as the ability to accommodate mineral deposition.
Although there are at least 18 isotypes of collagen, only a few have a clearly understood role in skeletal function and structure (Table 1.1). Type I is the main collagen in bone and has served as the scaffold in tissue-engineered grafts in clinical practice (see Appendix VII). Type II is associated with cartilage, type X with hypertrophic cartilage, and types III and V with granulation tissue. Types IV and VI play a role in the endothelial matrix. Fracture repair best elucidates the transitional evolution of collagen types in bone biology with early fracture collagens types II and III ultimately giving way to type I. In fact, aberrations in type III collagen production may result in nonunions or delayed unions (3).
Type I collagen contains two α(I) chains and a similar but genetically different α2(I) chain. Alpha chains consist of repeating triplets of glycine and usually proline and hydroxyproline. Posttranslational alterations include the formation of cross-links, glycosylation of lysyl or hydroxylysyl residues with glucose or galactose, and hydroxylation of lysyl residues.
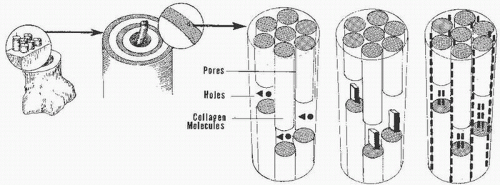
Type I collagen is an aggregate of fibrillary molecules that, by ultrastructural examination, demonstrate cross-striations (Fig. 1.3) (4). The axial periodicity is explained by a staggering or overlapping of four helicostructures. The initial deposition of mineral in bone most likely occurs in the gap regions of this fibrillary structure (Fig. 1.4).
Although several dozen types have been described, the relatively general term collagen encompasses different molecules of the extracellular matrix that share the triple helix characteristics described but also contain diverse properties from a clinical point of view. In fact, collagen can be divided into fibrillary (or interstitial collagen) and those considered nonfibrillary collagens. Nonfibrillary collagens, with the exception of the growth plate or periosteum, are not present in mineralized bone matrix.
Fibrillary collagens are more ubiquitous (3) (Table 1.1). The exact amino acid sequence of a collagen fibril may dictate certain physical and chemical properties such as its ability to interact with other components of the matrix and its very ability to form mineral. Bone collagen is biochemically almost pure type I, with a small component of type V. The relative absence of type III and other nonfibrillary collagens contributes to the ability of the matrix to mineralize. Of particular interest here is type X collagen, which appears to be a growth plate-specific collagen associated with endochondral bone formation both in the growth plate and in fracture repair. Expressed by hypertrophic chondrocytes, type X collagen is important in the remodeling of cartilage destined to be mineralized. Type IX collagen is associated with type II collagen, suggesting complex interaction between the two types of collagens, particularly with acetic elements of the cartilage matrix glycosaminoglycans. Type XII collagen, a component of embryonic tendon and periodontal ligament, may serve important functions linking fibrils and other matrix elements. Type II collagen is a marker of differentiated chondrocytes.
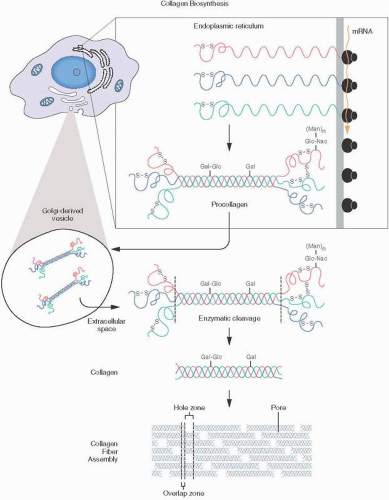
The synthesis of all collagens is complex and occurs via a multienzymatic biosynthetic pathway (Fig. 1.2). Deficiencies in certain parts of the pathway may be associated with hereditable disorders such as osteogenesis imperfecta, Ehlers-Danlos syndrome, and other conditions. In the nucleus of cells, DNA codes genes by transcription, splicing, and other processes. Messenger RNA leads to the assembly of pre-pro-α-chains in the polyribosomes of the rough endoplasmic reticulum. Chain association and disulfide bond formation occur in the Golgi apparatus, with eventual formation of the procollagen triple helix. Procollagen is extruded in secretory vesicles into the extracellular space, where enzymatic cleavages occur, leading to collagen formation. Fibril formation and lysyl oxidation lead to cross-linking and the resultant ultrastructurally detectable collagen. The transcription and the translation of the collagen genes are similar to those of other secretory proteins, which, possessing a very hydrophobic signal, cause the ribosomes involved in collagen synthesis to attach to the endoplasmic reticulum membrane and subsequently secrete the protein within the lumen of this organelle.
FIGURE 1.2. Collagen biosynthesis: messenger RNA is produced within the nucleus. Subsequent synthesis of collagen α-chains takes place in the endoplasmic reticulum. Key steps are the hydroxylation of lysine residues, development of disulfide bridges, and formation of triple helical procollagen molecules. After transport to the Golgi apparatus and secretion to the cell surface in Golgi-derived vesicles, procollagen is discharged into the extracellular space, where cleavage occurs, transforming the procollagen into tropocollagen, which then aggregates into collagen fibrils.
TABLE 1.1 Major Collagens in Bone and Cartilage
Collagen Type
Functions
Diseases
Type I
Most abundant collagen in bone matrix (90% of organic bone matrix) Also found in skin, tendon, and dentin
Osteogenesis imperfecta (oim mouse)
Type III Type V
Present in granulation tissue, fracture callus vessels (III), and synovial membranes (V)
Ehlers-Danlos syndrome
Type II
Major collagen in cartilage and perichondrium
Type X
Found in hypertrophic cartilage and calcified cartilage zone
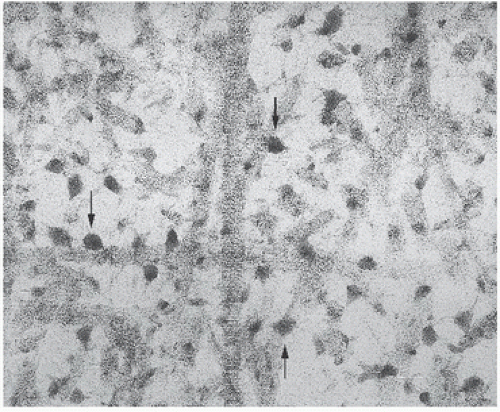
FIGURE 1.4. Collagen fibrils with enmeshed hydroxyapatite crystals (arrows) (ultrastructure).
The intracellular events are complicated, involve many different enzymes, and consist of hydroxylations and glycosylations. The three procollagen chains are stabilized by disulfide bridges. The cross-links pyridium and deoxypyridinoline, which are formed during collagen synthesis, are secreted in the urine and form the basis of the urinary analysis of bone turnover (5).
Collagen mutations associated with clinical entities or syndromes include osteogenesis imperfecta, or brittle bone disease. In some types of osteogenesis imperfecta, a basic type of defect involving type I collagen has been described. Structural mutations of type I collagen have been linked to the development of Ehlers-Danlos syndrome, type VII. Other mutations shown to influence collagen synthesis are at the transcriptional, translational, and posttranslational levels. Mutations such as those in the Ehlers-Danlos syndrome, type VII, which interfere with tensile strength, have little effect on bone, whereas other mutations, affecting primarily the folding of the triple helix, seem to have a dramatic effect on demineralized tissue (6).
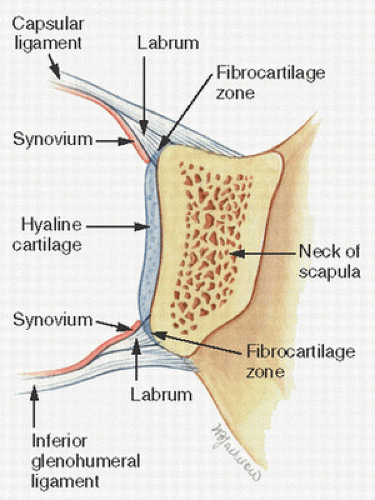
FIGURE 1.5. Fibrous tissue forms the basis of many osteoarticular connections, as seen in this wedge-shaped tissue attachment of the glenohumeral capsule and ligament, so-called glenoid labrum.
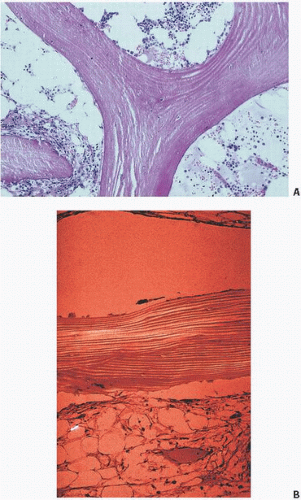
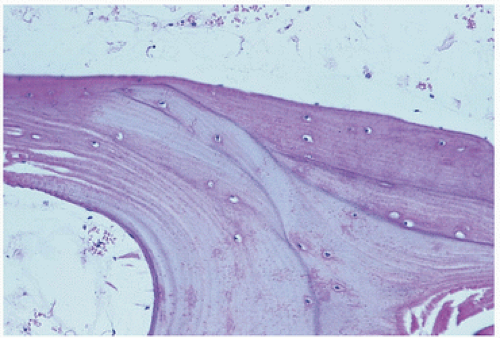
FIGURE 1.6. (A) Trabecular bone showing the normal organized pleated “lamellar” appearance. Note that cracking of bone induced by tissue processing occurs along lamellar collagen lines. (B) Trabecular bone with normal lamellar appearance (polarized light).
Cross-linking of collagen is critical for the physical and structural stability of extracellular matrix. Abnormalities of cross-linking have been associated with lathyrism, osteogenesis imperfecta, and other conditions.
Postulated hormonal interference with collagen synthesis includes inhibition of type I collagen synthesis in osteoblasts by 1,25-dihydroxyvitamin D3, PTH, interleukin 1 (IL-1), and glucocorticoids. Insulin-like growth factors (IGFs) and transforming growth factor beta (TGF-β) probably stimulate type I collagen.
Bone collagen is broken down in osteoclastic lacunae by MMPs (such as collagenase and cysteine proteases) and other mechanisms such as acidosis.
Large aggregation of collagen bundles may be discernible on light microscopy as successive layers of organized collagen, with these bundles consisting of finer units of highly arranged collagen fibers. Collagen fibers may be separated into those that are an intrinsic component of the bone tissue and those that are extrinsic to the bone proper, serving essentially a supporting anchoring function. The ligamentous insertions into the labrum of the shoulder glenoid labrum or the collateral ligaments of the knee are such examples (Fig. 1.5).
In normal mature bone, the intrinsic collagen fibers have an organized parallel arrangement (Fig. 1.6). In its most organized state, these parallel fibers appear as a stacking of successive, thin layers of parallel collagen deposition termed lamellae. Most evident under polarized light microscopy, these closely packed collagen fibrils run parallel but in different directions, one lamella to another—an organization similar to that of plywood. Numerous variations of this plywood lamella fibril organization can be seen, depending on the directions of the consecutive lamellae or rotational organizations, the optical qualities of which will differ accordingly (7).
When the fibrillary extracellular collagen matrix is more loosely packed and distributed without any specific spatial organization, a different appearance is noted, often referred to as woven or fibered bone matrix (Fig. 1.7). This is most usually characterized by a bone matrix that contains randomly distributed collagen and osteocytes. This matrix is often hypercellular and darker staining, may show increased mineralization, and is often associated with a higher rate of bone formation. Characteristic conditions in which this woven bone may be seen are the bone of the embryonic skeleton, bone produced by tumors, and bone associated with metabolic bone conditions of high metabolic activity such as Paget’s disease and fracture callus.
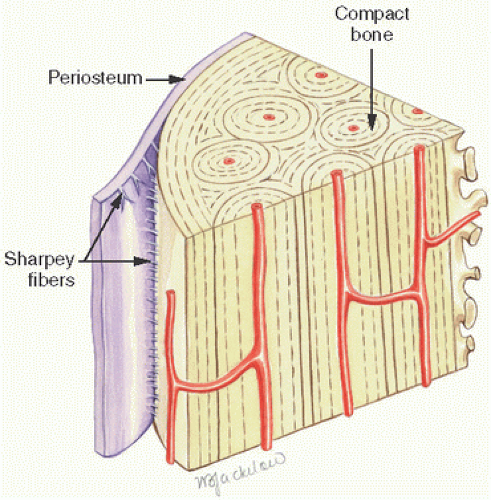
Extrinsic collagen fibers refer to those that correspond to the attachment process of soft tissues such as that of skin, tendons, ligaments, periosteum, muscles, and other tissues. Named by Collica in 1889, these are often referred to as Sharpey fibers (Fig. 1.8). These fibers are tightly packed collagen fibers that insert, often perpendicularly, into the intrinsic collagen fibers of the underlying bone. In essence, Sharpey fibers anchor the periosteum to the bone and may be mineralized at the fiber-bone interface. Weakness or developmental errors in the development of Sharpey fibers may result in clinical entities such as avulsion of the tibial spine in childhood or Osgood-Schlatter disease.
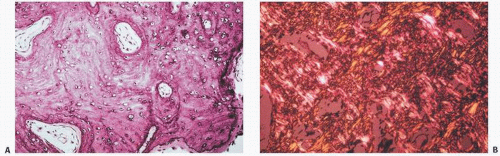
FIGURE 1.7. (A) Woven bone with its characteristic increased haphazard cellularity. There is irregularity to the osteocyte spatial distribution and increased numbers and sizes of osteocyte lacunar spaces. (B) Woven bone under polarized light demonstrating the loss of lamellar architecture.
FIGURE 1.8. Sharpey fibers are the tight collagen bundles that link soft tissues such as skin, muscle, tendons, and ligaments to mineralized tissue. Here Sharpey fibers are seen as radiating fibrous connections between cortical bone and the periosteum.
TABLE 1.2 Bone Composition
Minerals
50%-70%
Elements
Ca
34 wt%
P
16 wt%
Na
1-10 wt%
Fe, Mg, K, Al, Cu, Sr
<1 wt%
Ag, Ba, Cr, Pb, Sn, Mb, Si, Ni
<0.1 wt%
Organic matrix
20%-40%
Water
10%
Lipids
<3%
Other disturbances of these extrinsic fiber connections manifest in a broad range of clinically diverse musculoskeletal and rheumatologic disorders termed as enthesopathies.
Apatite
Bone mineral is essentially a calcium- and hydroxide-deficient analog of geologic mineral and is referred to as apatite or hydroxyapatite, Ca5(PO4)3OH (2). Apatite crystals are small. Because they contain impurities such as carbonate, magnesium, and acid phosphatase and are missing some OH, bone mineral is often termed poorly crystalline. However, bone crystals are more soluble than natural types of geologic crystals and thus can better capture and store calcium, phosphate, and magnesium. Naturally occurring geologic sources of apatite continue to serve clinical function with their application in bone grafting techniques (see Appendix VII) (Table 1.2).
Hydroxyapatite (or apatite) constitutes at least 90 percent of the total mineral weight. The mineral in other tissues such as calcified cartilage or primary spongiosa, cementum, dentum, and enamel also contain apatite, most likely of a different size and degree of perfection as indicated by biochemical studies that have examined calcium-to-phosphorus content, carbonate content, solubility, and x-ray diffraction patterns. Initial mineralized components may be different, including amorphous tricalcium phosphates and brushites, perhaps acting as precursors to hydroxyapatite. More than likely, the earliest mineral deposited in the bone extracellular matrix of collagen is a less perfect hydroxyapatite.
Mineralization
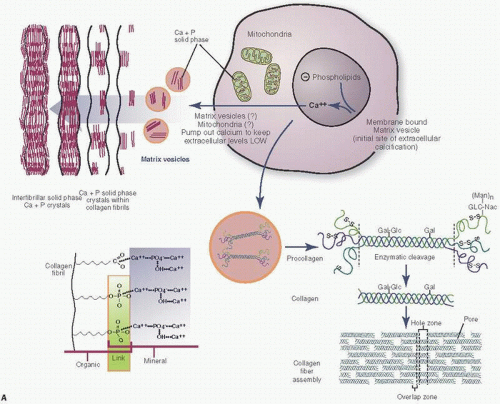
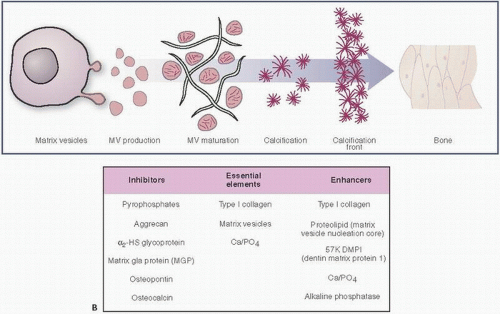
Mineralization, like blood clotting, is a phenomenon that requires a cascade of events. Promoters and inhibitors need to be finely balanced to result in functional bone. Initially, matrix vesicles develop in chondrocyte and osteoblast cells (Fig. 1.9). These vesicles, roughly 100 nm in diameter, contain calcium and inorganic phosphate ions (Pi), and initiate hydroxyapatite crystal formation, the building blocks of bone mineralization (8). Inorganic pyrophosphate (PPi) inhibits the ability of Pi to promote crystallization. Thus, in this tightly titrated milieu, the proper balance between the levels of Pi and PPi must be maintained. Calcium pyrophosphate crystal deposition disease (pseudogout) (see Chapter 15) is just one clinical example of a failure of this balance.
Eventually, the matrix vesicles reach the cell surface, form blebs, and initially protrude and eventually extrude into the extracellular environment. This exposure to the extracellular environment leads to further crystal growth and proliferation. Sulfated and/or phosphorylated proteins act as stimulators or “nucleators” of crystal growth. Access to calcium, phosphorous, and OH concentrations is required for the process to proceed. The role of alkaline phosphatase is less clear, but is associated with mineralization.
FIGURE 1.9. (A) The production of collagen and initiation of calcification by chondrocyte and osteoblast cells. (Continued)
Matrix vessels also include MMPs, which are responsible for matrix breakdown and turnover. For example, MMP-13, produced by hypertrophic chondrocytes, is secreted in matrix vesicles.
The accumulation of calcium in matrix vesicles depends, in part at least, on annexins, a family of calcium channel molecules, of which annexins II, V, and VI within the lipid bilayer of matrix vesicles are essential for calcium accumulation (9).
Mineralization Front (Calcification Front)
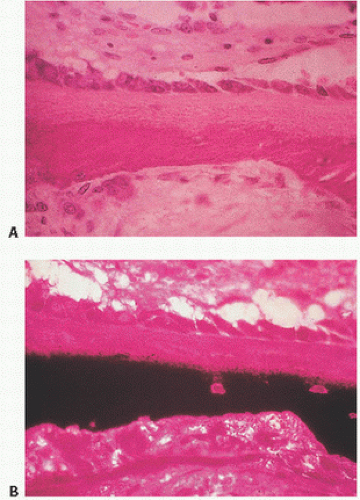
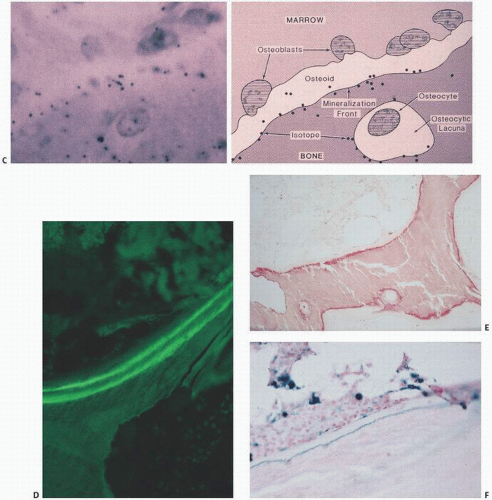
During bone formation, osteoblasts produce the collagenized protein that, under the normal sequence of events, will become bone. This tissue is referred to as osteoid. Osteoid is a composite of collagen and bone-forming proteins that, in the normal sequence of events, will undergo mineralization. In the normal state, approximately 20 percent of bone surfaces are covered by osteoid, which is usually approximately 10 µm in thickness and constitutes roughly 2 percent of structural bone (Fig. 1.10). It is identified as the layer of collagen deposited by osteoblasts. Osteoid begins to calcify about 10 days after its deposition, the zone of deposition identifiable on routine histologic sections as a basophilic linear line referred to as the mineralization or calcification front (Fig. 1.11). It is thought that the mineral replaces water, which occupies approximately half of the osteoid volume. Other terms for this are the French term ligne frontiere, demarcation line, or phosphate ridge. It is similar to the zone of calcification in cartilage referred to as the tidemark, which identifies the junction between calcified cartilage and normal articular cartilage in the articular bone. The changes that occur at the mineralization front are not well understood but may involve alterations in collagen or in the quantity and type of noncollagenized proteins. The mineralization front has significant clinical relevance: It is the site of deposition of bone-scanning agents (technetium diphosphonates), bone-labeling agents (tetracycline), and mineralization inhibitors such as aluminum (10).
FIGURE 1.9. (Continued) (B) Matrix vesicle calcification: inhibitors, essential elements, and enhancers. (Modified from Robey PG, Boskey AL. Extracellular matrix and biomineralization of bone. In: Favus MJ, ed. Primer on the Metabolic Bone Diseases and Disorders of Metabolism. 6th ed. Washington, DC: American Society of Bone and Mineral Research; 2006:12-19.)
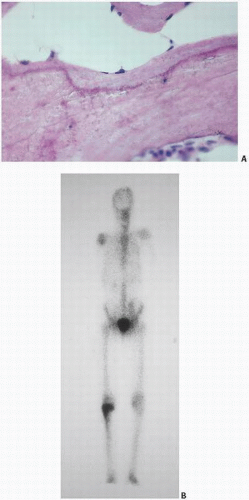
In clinical bone scanning, there are two reasons for a “positive” bone scan. First, the bone-scanning agent, usually technetium methylene diphosphonate, goes to areas with increased vascularity. Second, the technetium later resides in zones that are undergoing active mineralization (i.e., the mineralization front). Autoradiographic techniques have localized technetium methylene diphosphonate scanning agents to the active site of mineralization (Fig. 1.11). Thus, a bone scan is hot because active mineralization is taking place. This localization is very sensitive, albeit nonspecific. In the most widely applied bone-labeling studies in research, this phenomenon is exploited. Tetracycline binds at the mineralization front, most likely by attaching to the hydroxyapatite crystals. In fact, many types of bone-labeling agents have been used historically, making use of this phenomenon. Clearly, tetracycline goes where there is active mineralization, and its ability to do this has allowed for the determination of bone calcification and bone formation rates (see Chapter 3).
Indeed, in certain clinical states in which there is abundant availability of iron (such as in thalassemia) or aluminum (as in the excessive use of phosphate-binding aluminum gels), these substances attach at the mineralization front and cause pathologic conditions (Fig. 1.11). This has been most well established with aluminum-induced osteomalacia of renal-related bone disease, but cases of iron-induced osteomalacia have also been documented (11). There are many other clinical examples of the importance of the mineralization front in metabolic activity. In the treatment of osteoporosis with fluoride, new bone at the site of fluoride deposition is the result, most likely by the formation of a less soluble fluoride apatite compound. The fluoride has been visually identified histopathologically by the identification of a peculiar basophilic line at the mineralization front and around osteocytic lacunar spaces (12). In fact, fluoride given without concomitant calcium in therapy for osteoporosis may produce a frank hyperosteoidosis, mimicking osteomalacia.
FIGURE 1.10. Osteoid is the type I collagen matrix deposited by osteoblasts rich in bone-forming proteins, which, in the normal sequence of events, will become mineralized. Normally, about 20 percent of trabecular surfaces are covered by osteoid, and osteoid constitutes 2 percent of bone volume. Undecalcified section (A) hematoxylin-eosin; (B) von Kossa stain.
FIGURE 1.11. (A) The mineralization front: between osteoid (top) and mineralized bone (bottom), routine histologic sections of trabecular bone demonstrate a dark line (hematoxylin-eosin stain). (B) Bone scan with uptake (hot spot) in the right proximal tibia. Bone-scanning agents such as technetium seek mineralizing tissue and deposit at the mineralization fronts. (Continued)
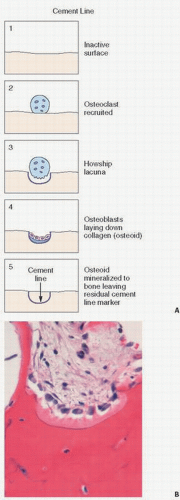
Cement Lines
Cement lines are distinct, usually basophilic staining regions of the cortical and trabecular bone matrix, which demarcate where bone resorption was completed and bone formation began (13) (Fig. 1.12). In hematoxylin-eosin staining, they usually are identified as a basophilic-appearing line, which usually has a smooth linear contour. In the classic understanding of bone remodeling, a bone formation unit (BFU) occurs when bone formation follows bone resorption. The cement line or reversal line in this situation indicates the period when bone formation begins following bone resorption (Fig. 1.13). In addition, cement lines may appear as arrest lines, which are thought to occur at sites where bone formation begins after a period of arrest. Cement lines provide evidence of previous times of bone formation and remodeling now buried within the matrix of cortical and trabecular bone. Numerous stains have been used to identify these cement lines, but routine hematoxylin-eosin sections not overly demineralized will reveal them.
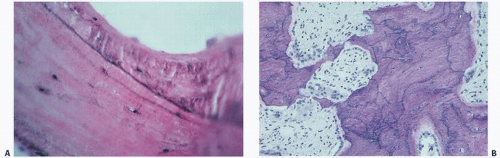
Cement lines in which there is activated or irregular bone remodeling, such as Paget’s disease, are characteristically markedly irregular, with a wavy or curly form pattern indicative of the woven bone appearance of these atypical bone states (Fig. 1.14). They are prominent in bone treated with fluoride and are particularly effective in demarcating the new fluoride bone from its host bone (Fig. 1.14).
FIGURE 1.11. (Continued) (C) Autoisotope study showing diagrammatically (right) and in vivo (left) the localization of previously injected technetium methylene diphosphonate along the mineralization front. (D) Tetracycline labeling (green) of bone (oral ingestion of two doses of tetracycline separated in time) reveals uptake at mineralization fronts and forms the basis of calculating mineralization rates (undecalcified section). (E) Aluminum (red) in dialysis osteomalacia localizes at the mineralization front blocking mineralization (undecalcified section, aluminum stain). (F) Iron (blue) in transfusional hemosiderosis or hemochromatosis localizes at the mineralization front (undecalcified section, Prussian blue stain).
FIGURE 1.12. Cement lines. Smooth basophilic lines within trabecular bone outlining previous units of bone that have been formed.
FIGURE 1.13. (A) Schematic representation of the formation of cement lines in bone. Osteoclasts are recruited and resorb a pocket within the bone (Howship lacunae). Osteoclasts disappear and are replaced by collagen-producing osteoblasts, which fill in the resorbed space. The collagen (osteoid) is subsequently mineralized, leaving the residual line at which previous resorption had ceased. (B) Histology of cemented line.
Resting Lines
Resting cement lines are those linear areas of basophilia on previously unresorbed bone surfaces where osteogenesis is thought to rebegin after a period of local cessation of bone deposition. Therefore, they have a more even appearance than those of the reversal cement lines. In general, cement lines are 1 to 2 µm in thickness and correspond to regions of mechanical weakness in bone (14). They may, in fact, be the site of facility of perpetuation of fracture lines.
The microstructure of cement lines and their composition are not very well known. Ultrastructurally, they appear as irregular granular dark structures with electron opacity. Collagen is probably scarce in the cement line, although lines in human bone are probably less mineralized than the surrounding bone or interstitial lamellae. Calcium-to-phosphorus ratios are significantly greater, however, than other parts of bone.
Histochemically, cement lines have been shown to contain an extrafibrillary matrix consisting of glycoproteins, proteoglycans, and phospholipids. Cement lines have also been shown to be highly species specific, with differences between cortical and trabecular bone, and woven and lamella bone. In fact, the number of cement lines may vary with age (15).
Bone-Specific Proteins
Noncollagenous Proteins
In addition to minerals and the extracellular collagen matrix, important components of bone are the noncollagenous proteins. Boskey and Robey (2) have classified them into four general groups (Table 1.3):
Proteoglycans, leucine-rich repeat proteins (LRPs), and hyaluronan
Glycosylated proteins
Mediators of cell attachment
δ-Carboxylic acid-containing proteins
Proteoglycans/Aggrecan
Aggrecan, the “aggregating” cartilage proteoglycan, is the main proteoglycan molecule in the cartilage matrix and is essentially responsible for providing the osmotic properties necessary for cartilage to resist compressive loads.
FIGURE 1.14. Prominent cement lines are seen in numerous conditions. In fluoride-treated osteoporotic bone, a prominent cement line separates normal (below) from fluoride-induced bone (above) (A). Irregular wavy, haphazard cement lines characterize Paget’s disease (B).
Collagenases that degrade interstitial collagen types I, II, and III
2 and 9
Degrade type IV collagen and gelatin
3 and 10
Stromelysins that degrade proteoglycans, laminins, fibronectin, gelatin, and casein
Proteoglycans are large molecules that contain glycosaminoglycans (acidic polysaccharide side chains) attached to a central core protein (16). Hyaluronan, a glycosaminoglycan not attached to a protein core, and versican, a large chondroitin sulfate proteoglycan, outline a territorial domain that may be destined for bone formation. Small leucine-rich proteoglycans (SLRPs or LRPs) such as decorin and biglycan are also present. Decorin is thought to cross-bridge or cross-link collagen fibers and regulate collagen fiber assembly. Mice without SLRPs contain large and more irregular collagen fibers, and those without biglycan have abnormalities in trabecular bone (17).
SLRPs may also bind TGF-β, which is important in cartilage development. Other SLRPs such as chondroadherin binds tightly to collagen and the α2β1 integrin. Double-knockout fibromodulin and biglycan animal models have been associated with joint laxity and supernumerary sesamoid bones.
Glycoproteins
This group includes osteonectin, the most abundant noncollagenous protein produced by bone cells, and alkaline phosphatase.
Osteonectin
Osteonectin, a glycophosphoprotein, is a noncollagenized protein identified in 1981 by Termine et al. (18). It avidly binds collagen and apatite. It inhibits hydroxyapatite crystal growth in part by binding calcium, but, in addition, binds type I collagen, the latter perhaps increasing the binding of apatite to the collagen and in effect stimulating nucleation under proper circumstances. In fact, there may be numerous bone proteins similar to osteonectin belying the complex interaction and stimulation of these biochemical events.
Although initially discovered in fetal bovine bone, it has subsequently been identified in adult bovine bone and fetal and adult human bone.
Osteonectin activity is much higher in trabecular bone, which is opposite to what is found with osteocalcin. Its activity has been noted in osteoblasts, young osteocytes, osteoid bone matrix, chondrocytes, and the mineralized matrix of chondroid bone, but not nonmineralizing cartilaginous cells. It has also been seen in cells with high turnover that are nonmineralized, such as the gut epithelium, and most likely is associated with extracellular matrix formation, especially that of bone and basement membrane. It has also been seen in association with certain platelets.
As a specific marker of metabolic function in the skeleton, osteonectin is more puzzling than that of osteocalcin. Therefore, its function is still speculative.
As with osteocalcin, osteonectin’s use in diagnostic pathology has been explored. Serra et al. (19) found strong positivity in osteosarcoma and osteoblastoma, and weaker staining in chondrosarcoma, Ewing fibrosarcoma, malignant fibrous histiocytoma, and brown tumors of hyperparathyroidism.
Alkaline Phosphatase
Alkaline phosphatases are a group of glycoproteins found in liver, bone, kidney, intestine, and placenta. The isoenzymes from liver, bone, and kidney are from the same gene product, but differ on the basis of heat and urea stability and electrophoretic mobility. Localized to the outer plasma membrane of osteoblasts, the role of bone-specific alkaline phosphatase remains somewhat enigmatic despite the fact that its deficiency in the clinical disorder hypophosphatasia results in dramatic skeletal abnormalities (see Chapter 3).
Mediators of Cell Attachment
Cell-surface receptors such as integrins mediate cell attachments utilizing many proteins, including type I collagen, fibronectin, thrombospondins (including oligomeric protein of COMP), vitronectin, fibrillin, BAG-75, osteoadherin, and members of the N-glycosylated protein (SIBLING) family, which include osteopontin and bone sialoprotein (2). All of these contain the cell attachment sequence (RGD or Arg-Gly-Asn), which allows binding to the integrin class of cell-surface molecules. Of the group, osteopontin and bone sialoprotein are known to anchor osteoclasts to bone.
Osteopontin is an acidic calcium-binding glycophosphoprotein found in the extracellular matrix and body fluids. Also referred to as pp69, Eta-1, 2ar, 2B7, and 44-kDa bone phosphoprotein, it is a cell adhesion protein and cytokine for CD44 and several integrins, and has been shown to be involved in dystrophic calcification, oncogenesis, and macrophage regulation. High levels of osteopontin have been correlated with tumor invasion, progression, and metastases (20).
COMP, a large glycoprotein that is abundant in cartilage, organizes collagen molecules closely spaced in parallel, resulting in rapid and efficient collagen fibril assembly. It also functions to maintain collagen network stability by binding to globular domains of collagen type IX. Composed of five flexible arms each terminating in a large globular domain, COMP structure has been likened to a bouquet of flowers (9). Mutation in COMP results in two skeletal dysplasias:
EDM1, a form of multiple epiphyseal dysplasia
Pseudoachondroplasia (PSACH)
The mutations initiate irregular folding of intracellular COMP, resulting in retention of the protein in the rough endoplasmic reticulum of chondrocytes (21). The gene coding for COMP maps to chromosome 19p13.1.
δ-Carboxylic Acid-Containing Proteins
δ-Carboxylic acid-containing proteins of note include osteocalcin (also known as bone gla protein or BGP) and matrix δ-carboxylic acid (also known as matrix gla protein or MGP). Whereas MGP is more ubiquitous in connective tissues, osteocalcin is more bone-and dentin specific. Excessive calcification in bone and soft tissue is the net result in MGP-deficient mice.
Osteocalcin: γ-Carboxyglutamic Acid Protein
Osteocalcin is a calcium-binding matrix protein made by osteoblasts. Its calcium affinity is related to vitamin K and γ-carboxyglutamic acid (GLA) residues.
Vitamin K is an essential cofactor for the formation of GLA residues in certain proteins (22). All GLA-containing proteins have a high affinity for binding calcium. In humans, there are two distinct groups, one of which predominantly occurs in calcified tissues. These include osteocalcin found in bone and a plaque GLA protein, which has been identified in the calcified vessel walls of atherosclerotic plaques. The GLA-containing proteins found in calcified tissues help regulate calcium deposition in these tissues, perhaps by strongly inhibiting the precipitation of calcium salts and hydroxyapatite growth. In fact, women with vitamin K antagonists during the first trimester of pregnancy may be associated with bone defects in their fetuses (23).
Osteocalcin constitutes 20 percent of the noncollagenous proteins of bone matrix. It has been isolated from more than a dozen vertebrate species and has been identified as a mature protein containing approximately 50 amino acids in a single peptide chain, with molecular weight ranging from 5,200 to 5,900 Da. It binds weakly to free calcium ions, but more strongly to calcium and hydroxyapatite crystals, and probably undergoes conformational shifts when associated with the apatite crystal as a result of electrostatic attraction between specific and ionic residues in the proteins and the calcium ions embedded in the face of the crystal. Its biologic properties are linked to this unique structure, and its affinity for apatite most likely affects bone mineralization directly. Osteocalcin is an effector molecule for 1,25-hydroxy D, which paradoxically stimulates osteocalcin synthesis but depresses collagen synthesis in the expression of alkaline phosphatase activity in cell proliferation.
Although found in the serum and therefore useful as a marker for certain metabolic bone diseases, most osteocalcin is incorporated into bone and has a half-life of approximately 5 minutes. It is metabolized in the renal tubule and has a circadian rhythm peaking at 1400 hours, a rhythm that may be adversely affected by glucocorticoids or growth hormones. In clinical situations, it has been extensively studied. It is produced by osteoblasts, not osteoclasts, and is concentrated in osteocytes. It may play a role in osteoclast recruitment but its release into the circulation most likely signals the cascade of bone turnover. Close correlations have been noted between osteocalcin and alkaline phosphatase activity and histomorphometric studies of bone mineralization. As with alkaline phosphatase, it increases in growth spurts and is particularly associated with puberty.
Osteocalcin is associated with coagulopathies during pregnancy and with warfarin administration. It is low in certain forms of congenital hypoparathyroidism, the George syndrome, and rises rapidly in response to single injections of calcitriol. Osteocalcin has also been noted to be low in certain populations of postmenopausal osteoporotic women, most likely reflecting a subpopulation with low bone turnover osteoporosis. In fact, it has been used as a marker for increase in bone formation as a response to fluoride therapy in the treatment of postmenopausal osteoporosis. Cooper et al. (24) reported decreased osteocalcin levels in 41 elderly women with hip fractures. It is interesting to note its activity with regard to estrogen therapy. Estrogen is thought to slow the rate of bone loss by reducing bone turnover, particularly the resorptive phase. Osteocalcin has been shown to increase following oophorectomy, diminish in response to estrogen replacement therapy, and rise again with cessation of hormonal therapy. Serum osteocalcin has been found elevated in all stages of Paget’s disease, including that in the initial osteolytic stage.
Osteocalcin has also been noted to be higher in the serum of patients with blastic and osteoblastic metastases compared with lytic metastases from the breast. It also reflects the high turnover in hyperthyroid states and has been noted to be decreased in patients with endogenous or exogenous glucocorticoid excess, conditions known to be associated with increased bone resorption.
Most recently, it has been used as a bone marker in diagnosing and differentiating bone tumors. Iwasaki et al. (25) have used it to differentiate osteosarcoma from chondrosarcoma, malignant fibrous histiocytomas, and giant-cell tumor.
In addition, there are serum-derived proteins largely composed of serum proteins such as albumin and α2-HS-glycoprotein, growth factors, and other molecules. α2-HS-glycoprotein-deficient mice develop ectopic calcification. Other components include water (˜10 percent of bone weight) and lipids.
Matrix Metalloproteinases
MMPs remodel the extracellular matrix (26). Present in bone, cartilage, ligaments, and tendons, the balance of MMPs and their inhibitors (tissue inhibitors of metalloproteinases or TIMPs) dictates a wide range of tissue alterations and disease states (Table 1.4). MMP-9 is a key identifiable MMP in bone degradation and MMPs 1, 3, and 13 in arthritic states. MMPs have also been incorporated into scaffold design to manage the process of degradation in engineered tissue products.
TABLE 1.4 Factors that Regulate Osteoclast Activity
Stimulation of osteoclast formation and/or resorption is denoted by (+); inhibition of formation and/or resorption is denoted by (-).
Modified after Roodman GA. Advances in bone biology: the osteoclast. Endocr Rev. 1996;17:308-332.
Bone Cells
In the development of an embryo, differentiation takes place along several cell lineages. Epithelial tissue develops from the ectoderm, mesoderm, or endoderm, whereas mesenchymal tissue develops from the mesoderm. Epithelial cells are characterized by close cell-to-cell contact with distinct ultrastructurally recognizable boundaries, so-called desmosomes and other junctions. On the other hand, mesenchymal cells have little or no contact with each other and, in general, are surrounded by an extracellular matrix, be it collagen, cartilaginous tissue, bone, or variations of the three. In the embryo, the cells that make up skeletal tissue (cartilage, bone, tendons ligaments, and marrow stroma) are thought to derive from a primitive and probable pluripotential mesenchymal stem cell.
A characteristic of mesenchymal tissue derived from the mesoderm or middle embryonic layer is the ability of the cells to migrate and occupy space between ectodermal and endodermal cell layers, an essential feature when considering the remarkable ability of the periosteum to respond to injury, the ability of the dermal tissue of the skin to close after a wound, and, in a larger sense, the phenomenon of all wound repair. Before identifying the individual characteristics of differentiated cells, at certain stages of development, several generalizations may be made concerning the mesenchymal stem cell (Fig. 1.15). First, mesenchymal stem cells, which in experimental models give rise to cartilage and bone in vivo, can be manipulated in vitro. Second, they have a linear progression with identifiable individual steps. Third, there is a local queuing phenomenon dependent on factors (or at least involving protein factors) that can be manipulated experimentally to alter size and function. Fourth, chondrocytes and osteoblasts, although stemming from the same primitive cell, after differentiation have a uniquely different appearance and function. Chondrogenesis, for example, requires the absence of vasculature, whereas osteogenesis is dependent on the presence of a vascular supply.
The pluripotentiality of mesenchymal cells is noted in the cells of the periosteum, a layer that responds dramatically to injury by forming woven bone. However, periosteal cells retain their developmental potential to be manipulated into two different tissues, bone and cartilage. For example, cultured periosteal cells implanted into porous ceramics in nude mice differentiate into osteoblasts, whereas the same tissue injected into subcutaneous sites in nude mice differentiates into both bone and cartilage.
Osteocytes
Osteocytes are the most abundant and longest lived bone cells. Their osteocytic canalicular system constitutes the most extensive bone compartment, the surface of which has been estimated to be 100× larger than the surface area available to osteoblasts and osteoclasts (27). The dense network of osteocytes ensures that no part of bone is more than several microns from a lacuna containing an osteocyte. Derived from the bone-forming osteoblast, osteocytes get embedded in the material they synthesize. Within bone matrix, they develop connections (dentritic cell processes) that connect the osteocytes to the bone surface.
Bone remodeling is thought to begin when microcracks alter the environment of the canalicular channels stimulating the osteocytes to translate mechanical strains into biochemical signals (see “Bone Remodeling”) (28).
The osteocyte network constitutes a huge mineral reservoir, but the physiology of this local mineral homeostasis remains largely enigmatic. Osteocytes are believed to control body phosphate levels, but their ability to resorb the pericellular bone (“osteocytic osteolysis”) remains a somewhat controversial concept.
It has been postulated that the bone remodeling signals produced by the osteocyte originate in the ability of the glycocalyx to form strong integrin attachments that may serve as mechanotransducers that transmit the mechanical signals to the osteocyte cell body, leading to the opening of hemichannels that allow for exchange of local tissue factors (28).
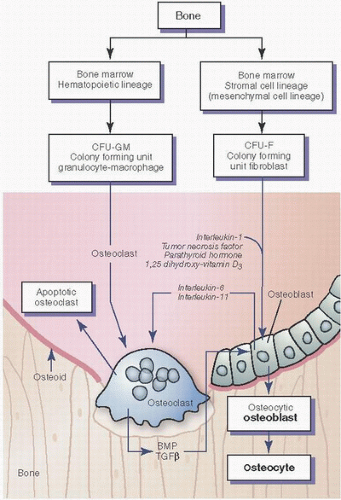
FIGURE 1.15. Origin and action of osteoclasts and osteoblasts. Osteoclasts arise from marrow hematopoietic cells, and osteoblasts from marrow stromal cells. Hormones such as parathormone and 1,25-vitamin D3, and cytokines such as IL-1 and TNF stimulate bone resorption in part by action on osteoblasts, which secrete their own cytokines such as IL-6 and -11. Thus, osteoclastic bone resorption is mediated through osteoblasts. Osteoclasts are not attracted to and will not adhere to osteoid, and therefore require degradation of osteoid to expose mineral. Osteoclastic resorption of bone releases bone matrix proteins and cytokines (such as TGF-β), which stimulate osteoblast production and bone formation. (Modified after Manolagas SC, Jilka RL. Mechanisms of diseases: bone marrow, cytokines and bone remodeling: emerging insights into the pathophysiology of osteoporosis. N Engl J Med. 1995;332:305-311.)
Osteocytes may also perceive and respond to canalicular fluid flow, piezoelectric fields, and streaming potentials. Reduced fluid flow causes apoptosis in vitro.
Osteocytes die by apoptosis, which can be accelerated in advancing age, in estrogen deficiency, during corticosteroid therapy, and after damage to bone. Older bone has a reduced number of osteocytes and accumulates microdamage.
There is evidence that osteocyte apoptosis is a key controlling step in the activation of osteoclastic resorption after fatigue microdamage, thus linking the osteocyte to initiation of the bone remodeling cycle (29). Osteoclasts, then activated, most likely dispose of both exposed bone and the apoptotic osteocyte.
Sclerostin, a WNT signaling molecule secreted mainly by osteocytes, functions as a potent inhibitor of osteoblast activity. Of note, sclerostin knockout mice have a high bone mass phenotype.
Pathologically, abnormal regulation of sclerostin has been observed in osteoporosis-related fractures, failure of implant osseous integration, metastatic bone disease, and select genetic diseases of bone mass such as sclerosteosis (30). In sclerosteosis, patients have increased bone mineral density and syndactyly due to sclerostin loss-of-function mutations. The sclerostin antibody (AMG 785; Amgen) decreases endogenous levels of sclerostin, allowing osteoblast survival and increased osteogenesis. Sclerostin most likely acts by inhibition of the WNT/β-catenin pathway in osteoblasts, binding LRP, the low-density lipoprotein receptor-related protein, and 4/5 receptor on the membrane of osteoblasts. Myeloma cells suppress osteoblasts by secreting sclerostin.
Ultrastructurally, the osteocyte resembles the osteoblast. It has a prominent nucleus and nucleolus. It also has abundant rough endoplasmic reticulum, well-defined areas of Golgi apparatus, mitochondria, microtubules, and microfilaments. With increasing mineralization engulfing the cell, the organelles are less readily discernible so that the osteocyte is identified by light microscopy only by its nucleus. Concurrent with this is a decreasing lacuna space.
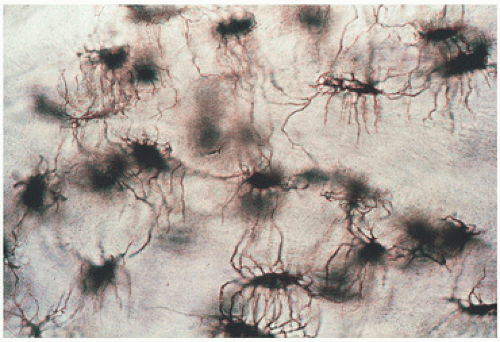
Osteocytes are in contact with osteoblasts on the bone surface by means of cell processes that travel through the canalicular system. Their ability to travel large distances is seen with special staining techniques (Figs. 1.16 and 1.17). These cell processes consist mostly of microfilaments, 5 to 7 nm in diameter, and may be detected to have met the processes of other cells by electron-dense side-to-side junctions, not dissimilar in some respects to the desmosomes of epithelial connections.
Osteoblasts and young osteocytes possess histochemically demonstrable alkaline and neutral phosphatase activity on their cell membranes. Because these phosphatases are located on cell surfaces, which also show binding affinities for positively charged molecules, the role of alkaline phosphatase in calcium transport is deemed important. The mature osteocyte, on the other hand, does not have membrane alkaline phosphatase to the extent it does when it is a young cell. The role of other enzymes such as 5·-nucleotidase is less clear, although it may affect intracellular cyclic AMP. Osteoblasts also contain adenylate cyclase. Cyclic AMP levels have been inversely correlated with collagen production by osteoblasts. These histochemical findings support a reduced metabolic activity of the osteocyte compared with the osteoblast.
FIGURE 1.16. Osteocytes: photomicrograph of osteocytes and osteocytic canaliculi seen by transmitted light in ground section (×10).
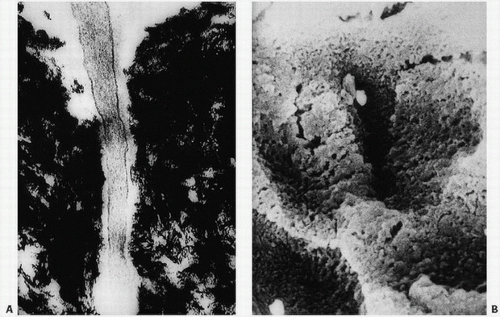
FIGURE 1.17. Osteocyte canaliculi: electron micrograph of a portion of an osteocytic process in an osteocytic canaliculus (A). In the scanning electron micrograph, the open space of the canaliculi is clearly evident (B).
Other histochemical studies have supported the role of the Golgi system in glycosylating procollagen prior to its secretion and its role in packaging the material into granules for secretion to the cell surface. Histochemical studies of osteoblasts and osteocytes have established a rich activity in osteoblasts, but sparse activity in osteocytes. Although this may reflect a lower metabolic activity of the osteocytic cell population, it may favor a longer life span for the osteocyte.
Although a role for the osteocyte in hormonal regulation of bone and resorption (osteocytic osteolysis) remains controversial, its extensive canalicular system suggests a function in transduction of mechanical loading.
Osteocytes may demonstrate plasticity, an activity with an ability to adapt to demand placed on them by manipulating its attachment to the bone matrix. Proteins such as collagen, integrin, and osteopontin may enhance matrix attachment, a process counterbalanced by producing digesting enzymes such as MMP-1.
Microcracks may also be a method of initiating remodeling. These microscopic cracks through bone may be either actual physical damage or osteocyte-induced apoptosis or necrosis, which sends signals for osteoblast recruitment.
Osteocytes express CD44, a transmembrane glycoprotein that binds fibronectin, collagen, and hyaluronate, a feature that may act as a marker for this cell. Osteocytes also express osteocalcin and galectin 2.
Proteins that identify the maturing osteocyte are:
E11
Alkaline phosphatase
Pi-regulating endopeptidase on chromosome (PHEX)
Matrix extracellular phosphoglycoprotein (MEPE)
Sclerostin
Fibroblast growth factor 23 (FGF-23).
Sclerostin and FGF-23 are produced by the osteocyte only after completion of differentiation. FGF-23 acts on the kidney to influence phosphate homeostasis (30).
Osteoblasts
Osteoblasts are bone cells of mesenchymal origin that synthesize type I collagen, the extracellular bone matrix, and noncollagenous proteins (Table 1.3). They arise from multipotential bone marrow stromal cells of the colony-forming unit of fibroblast lineage (CFU-F).
Osteoprogenitor cells, defined as cells capable of proliferating to give rise to progeny that can differentiate into connective tissue phenotypes such as bone, have been harvested in bone marrow aspirates from several bone sites, including the iliac crest, proximal aspect of the tibia, and even the calcaneus (31). Iliac crest bone appears to provide the highest yield.
Fully functional osteoblasts can be characterized as cells that express messenger RNA for a-1 procollagen, alkaline phosphatase, and bone-forming proteins such as osteopontin and osteocalcin (GLA protein).
The osteoblast has been viewed as a sophisticated fibroblast in that they are difficult to distinguish in cell culture. However, at least two osteoblast-specific transcripts have been identified that code for
A member of the runt family of Drosophila vertebrate homologs, Cbfa1 is critical for bone development and is one of the earliest and most specific markers of osteogenesis. Its deficiency in Cbfa1-deficient mice results in perinatal lethality, a skeleton consisting exclusively of cartilage. Mice heterozygous for it develop cleidocranial dysplasia, the most frequent human skeletal disorder.
RUNX2 is a “master” transcriptional regulator. Its roles include:
Activation and repression of phenotypic genes as osteoblasts progress through stages of differentiation.
Regulation of genes related to bone resorption and osteoclastic coupling, and
Mediating the transcriptional and functional activity of osteogenic signaling pathways (32).
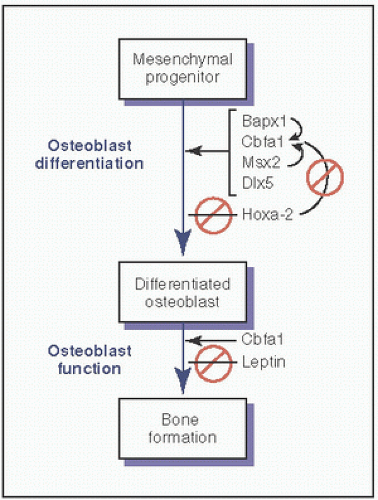
Additional homeobox proteins that affect osteoblast differentiation include Dlx5, Bapx1, Msx2, and LMP-1 (33) (Fig. 1.18).
Osteocalcin (OC)
Osteocalcin is produced by mature osteoblasts. Its synthesis is stimulated by 1,25-dihydroxyvitamin D. In the osteoblast, OC undergoes posttranslational modifications (γ-carboxylation) that require vitamin K1 (phylloquinone) as a cofactor. Carboxylated osteocalcin (cOC) has a high affinity for hydroxyapatite and mineral ions and is a determinant for calcium distribution in bone tissue. The majority of cOC accumulates in the bone matrix. A small fraction is also present in serum and is a sensitive marker of bone formation and a predictor of fragility fractures.
FIGURE 1.18. Key factors in osteoblast differentiation. Bapx1: a homeobox protein that promotes osteoblast differentiation; Cbfa1: a transcriptional regulator (also called RUNX2); Msx2: a homeodomain protein/transcriptional regulator that controls aspects of craniofacial development and is maximally expressed in mesenchymal/osteoprogenitor cells, but a negative regulator of some genes expressed in mature osteoblasts; DLX5: a homeodomain protein/transcriptional regulator that plays a role in limb development; HOXA2: a member of the vertebrate homeobox gene family that encodes transcription factors; leptin: a signaling molecule that is antiosteogenic in vivo, but in the treatment of leptin-deficient mice can stimulate chondrocytes and osteoblasts. (Modified from Lian JB, Gordon JA, Stein GS. Redefining the activity of a bone-specific transcription factor: novel insights for understanding bone formation. J Bone Miner Res. 2013;28(10):2060-2063.)
Undercarboxylated osteocalcin is thought to play a role in inhibiting extracellular matrix mineralization.
Only gold members can continue reading. Log In or Register to continue