Fig. 2.1
Examples of materials used clinically to treat wounds and experimental materials currently in research stages
Rather than a passive gel dressing that will have a minimal role in the healing process, technologies are needed that will actively promote wound healing by acting as a substrate for endogenous cell migration and proliferation. Cell-based therapies, such as the application of stem cells, may meet these needs.
2 A Brief Introduction to Stem Cells Derived from the Amniotic Fluid
The cell source used in cellular therapies for wound healing is an important consideration that has implications in the cost, speed, and outcome of the treatments. Human keratinocytes are perhaps the optimal cell type to employ as they make up the epidermal zone of skin tissue. However, autologous and allogeneic keratinocytes suffer from the same drawbacks as their autologous and allogeneic skin graft counterparts, i.e., secondary surgical sites and potential for rejection, respectively. This begs the question: Can we employ cells that are beneficial to wound healing, but may be immunoprivileged despite originating from an allogeneic source?
Mesenchymal stem cells (MSCs) have shown therapeutic potential for repair and regeneration of tissues damaged by injury or disease. In particular, MSC treatment of acute and chronic wounds result in accelerated wound closure, increased epithelialization, formation of granulation tissue and angiogenesis [14]. Amniotic fluid-derived stem (AFS) cells are a cell type that shares some characteristics with MSCs, but are perhaps even more potent as they are sourced from an earlier point along the developmental timeline, namely the gestational period. As such, they are an attractive cell source for applications in regenerative medicine due to their high proliferation capacity, multipotency, immunomodulatory activity, and the lack of significant immunogenicity [15, 16]. Unlike embryonic stem cells (ESCs), AFS cells do not form teratomas when injected into immune-deficient mice. Furthermore, AFS cells remain stable and show no signs of transformation in culture. The isolation of AFS cells is a simpler process than that for isolation of MSCs and large numbers of AFS cells can be isolated and expanded from as little as 2 mL of amniotic fluid. These cells proliferate rapidly with doubling times of 30–36 h, and do not require supportive feeder layers [15]. The immunomodulatory and high proliferation properties of AFS cells suggest that they may be appropriate as an effective “off the shelf” cell therapy product for wound healing.
3 Flexibility of Amniotic Fluid-Derived Stem Cells in Regenerative Medicine
In recent years AFS cell-based research for applications in regenerative medicine has become more prevalent in the literature. As with other stem cell types that were established earlier such as MSCs, adipose-derived stem cells (ADSCs), and ESCs, AFS cells have been explored for force induced differentiation into cells of various tissue types, for repairing and replacing damaged or diseased tissues, and antitumor treatments. Additionally, the secretory profiles of AFS cells suggest they may be effective as deliverable “drug factories,” efficiently secreting cytokines and growth factors at target sites such as MSCs have been shown to do [17].
For differentiation purposes, AFS cells have been force induced into a number of lineages, including those of the liver [18], pancreas [19], cartilage [20], fat, muscle, bone, neuronal, and endothelial lineages [15, 21]. In addition to a level of multipotency, AFS cells share several other characteristics with MSCs, including the ability secrete cytokines, many of which are pro-regenerative or immunomodulatory. The ability to secrete biological molecules such as these makes AFS cells, like MSCs, useful for various reparation therapies as well as antitumor treatments. The immunomodulatory activity of AFS cells has been implemented to treat chronic allograft vasculopathy, by decreasing inflammation, thus preventing intimal wall thickening and loss of the graft [22]. This immunomodulatory activity has been further shown in controlled in vitro immune reaction studies, in which AFS cells as well as cell-free supernatants from AFS cell cultures inhibit lymphocyte activation, suggesting the responsible mechanism is through secreted soluble immunomodulatory factors [16]. These characteristics, secretory profiles and low immunogenicity, have also served as motivation to explore the use of AFS cells in anticancer treatments. Like MSCs, AFS cells show a propensity to home to tumor sites with administered intravenously [23]. In one such example, AFS cells expressing interferon beta were administered systemically to mice in a bladder tumor model, where they homed to the tumors and succeeded in inhibiting tumor growth and prolonging survival [24].
This ability to secrete growth factors, immunomodulatory factors, anti-inflammatory molecules, and to migrate to sites of injury has suggested another use in cell therapy—namely can AFS cells be implemented in wound healing treatments in skin?
4 Perinatal Wound Healing
There is a vast amount of literature that argues towards the potential that AFS cells have in wound healing applications, which stems from the phenomenon of perinatal, or fetal, wound healing, in which injuries to a fetus can heal completely scar free [25]. During development the fetus is surrounded by amniotic fluid (AF) within the amniotic sac, the same environment from which AFS cells are isolated. Components of the AF may be responsible for its regenerative properties, and also be able to imbue the regenerative properties inherent in AFS cells. A number of studies have determined that the amniotic fluid has a unique composition that changes over time, supporting scar-free healing at the early stages of gestation, and then transitions to more traditional healing processes at the end of pregnancy. For example, human amniotic fluid was assessed in vitro for its ability to regulate several major proteases involved in wound healing. It was found that amniotic fluid enhanced collagenase, thereby inhibiting collagen formation, a large component of scarring. Conversely, activities of hyaluronidase were inhibited, thereby allowing hyaluronic acid (HA), an anti-inflammatory glycosaminoglycan common to the extracellular matrix (ECM), to be present in high levels [25]. Furthermore, it has been demonstrated that fibrotic healing that occurs in late gestational and adult wounds is correlated with increased hyaluronidase activity, which decreases HA levels [26]. HA has often been associated with regeneration and anti-inflammation, and is now commonly used in applications demanding such characteristics [27].
Amniotic fluid-based fetal wound healing studies have yielded a knowledgebase consisting of growth factors, cytokines, and extracellular matrix components that have been further explored to minimize scarring in adult wounds. Indeed, it has been shown that amniotic fluid, containing high levels of basic fibroblast growth factor (bFGF) and platelet-derived growth factor (PDGF), was a potent stimulant of both fetal and adult skin fibroblast growth [28]. A number of clinical trials have been instigated, but none have reached the level of FDA approval [29]. However, the concept of using the fetal developmental environment to “fool” the body into scar-fee healing holds merit.
5 Amniotic Fluid-Derived Stem Cells in Wound Healing
Secretory profiles of AFS cell-conditioned media contain a number of cytokines known to be important in wound healing, such as vascular endothelial growth factor (VEGF), epithelial growth factor (EGF), and transforming growth factor beta (TGF-ß) [30]. This suggests that AFS cells delivered to wound sites may be able to secrete these factors in situ, contributing to wound healing in cell therapy administrations. MSCs have been shown to have a paracrine role in wound healing, secreting similar factors [31]. Interestingly, MSCs have also been documented as being able to integrate permanently by differentiating into cells of myofibroblast phenotype in vivo [32] and acting as vasculature-supporting pericytes in vitro [33], another potential mechanism by which AFS cells could potentially impact wound healing.
As discussed above, the environment within the amniotic sac itself may be the main component in effective wound healing, but until recently it was unknown whether a cellular component of the environment was important. In a fetal lamb wound model, fetuses receive an infusion of autologous AFS cells into the amniotic sac. Access to wounds was either not impeded, or made impermeable to cells using exterior titanium chambers. AFS-impermeable wounds showed significantly slower healing rates. Furthermore elastin content in those wounds were significantly decreased compared to wounds that were accessible to the AFS cells, further indicating the effectiveness of the cells themselves in wound healing, albeit in a fetal environment [34].
Our laboratory has a track record in pioneering isolation, characterization, and applications of AFS cells [15, 19, 21, 35, 36]. However, until recently no one had explored the use of human AFS cells in healing of severe wounds in adults. We recently demonstrated the use of AFS cells using a full thickness wound model in mice [13]. We employed a bioprinting device previously developed in house [12] to deliver AFS cells to wounds within a fibrin-collagen gel vehicle, and compared this treatment to MSCs or no cells delivered in the same gel delivery vehicle. AFS cells and MSCs showed similar potency in topographical wound closure (Fig. 2.2), significantly accelerating wound closure compared to the cell-free control. However, upon histological inspection, we observed that AFS cells induced significantly greater vascularization than both MSC and cell-free treatments (Fig. 2.3a–h) during the short 2-week time course. Additionally, vasculature in AFS-treated wounds also appeared more mature, displaying robust smooth muscle actin surrounding larger vessels and a lack of extravasated red blood cells (Fig. 2.3i–n). Extravasated red blood cells were observed in the other groups, indicating leaky or less mature blood vessel structures. Interestingly, both AFS cells and MSCs were transient in our model as evidenced by tracking of the cells after transfection with green fluorescent protein (GFP). Cells were present immediately after treatment, 4 days after treatment, and 7 days after treatment—albeit in fewer numbers. However, no evidence of either cell type was found after day 7 (Fig. 2.4).
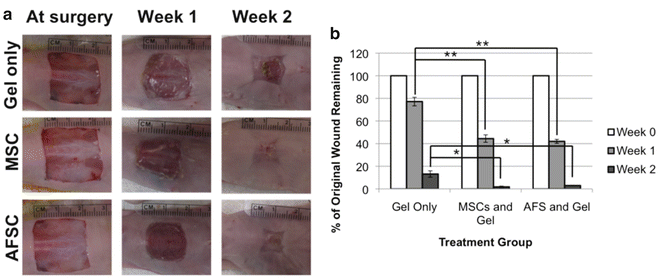
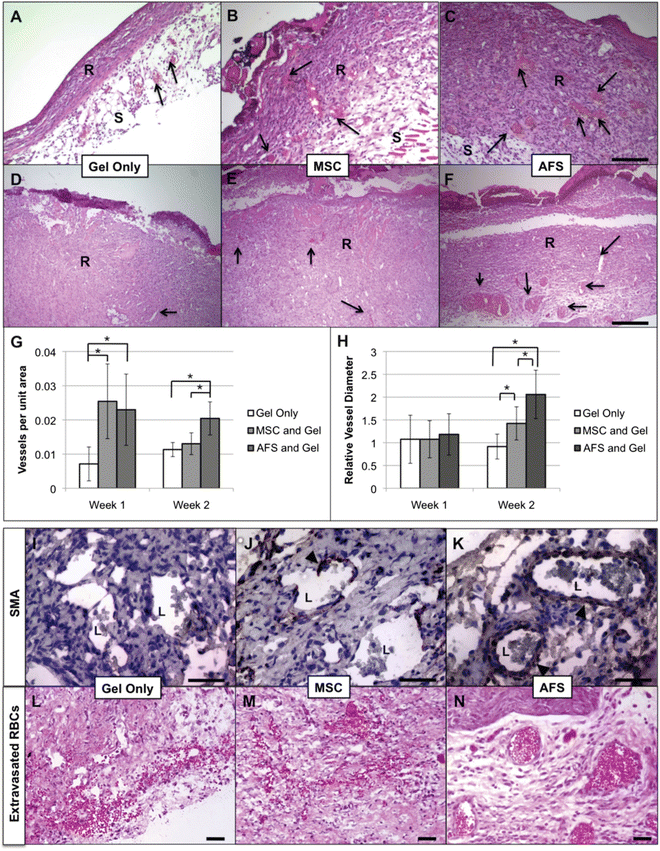
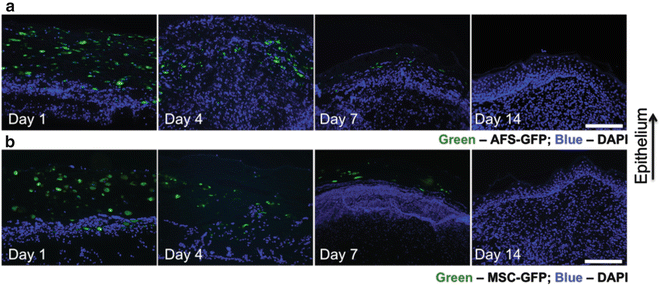
Fig. 2.2
Wound closure rates are increased in AFS- and MSC-treated mice. (a) Gross histology images illustrating wound closure in Gel-only, MSC, and AFS treatments. (b) Percentage of unclosed wound remaining at surgery, 1 week, and 2 weeks. Significance: *p < 0.05; **p < 0.01
Fig. 2.3
AFS cells induce neovascularization and blood vessel maturation in vivo. H&E staining revealed thicker regenerating tissue w/more blood vessels in AFS and MSC group compared to the gel-only group. (a–c) Week 1; (d–f) Week 2; (a and d) Gel-only; (b and e) MSC; (c and f) AFS. Arrows—vessels. AFS cells increase the number of newly formed vessels and induce formation of larger vessels. (g) Microvessel density (MVD) and (h) vessel diameter were quantified using histology images and ImageJ software. Significance: *p < 0.05. AFS cells induce formation of mature blood vessels. SMA staining around blood vessels in regenerated tissues of gel only (i), MSC (j), and AFS (k) groups. Arrows: Cells expressing SMA. Extravasated RBCs were present in gel only (l) and MSC (m) groups, but not in the AFS (n) group
Fig. 2.4
AFS cells and MSCs are transient in the regenerating wound and do not permanently integrate into the tissue. Regenerating skin was harvested at day 1, day 4, day 7, and day 14 in order to determine the presence of labeled cells. GFP-labeled AFS cells (a) and MSCs (b) are visible in decreasing numbers of the time course of the experiment. Green: GFP-expressing AFS or MSC; Blue: Nuclear staining, DAPI
Nevertheless, these results suggested that AFS cells have the potential to be an effective therapy for treating skin wounds. Based the transient nature of the cells, we hypothesized that the augmentation of skin regeneration was due to trophic delivery of cytokines. To further investigate this concept, we first generated conditioned media from these particular populations of AFS cells and MSCs, and analyzed it for a panel of potent growth factors using a proteomics array. As expected, samples tested positive for cytokines such as bFGF, VEGF, hepatocyte growth factor (HGF), and members of the insulin growth factor-binding protein (IGFBP) superfamily (Fig. 2.5). In general, AFS-conditioned media tested positive for more of these cytokines and at higher concentrations. Based on the increased angiogenic activity noted above, we then hypothesized that AFS cells were likely responsible for recruiting endothelial cells via secretion of these factors. We next performed a Transwell migration assay using human umbilical vein endothelial cells as the migrating population. In order to simulate the mechanically soft environment of the in vivo wound bed we employed a 2 kPa stiff substrate versus much stiffer traditional plastic substrates on which the AFS cells were seeded as the recruiting population. In addition, a cell-free control was used. A greater number of HUVECs migrated when exposed to the AFS cultures compared to the cell-free control. Interestingly, AFS cells cultured on the softer 2 kPa substrate were significantly more adept at HUVEC recruitment, suggesting that the environment in which they exist plays an important role in their therapeutic effectiveness [13].
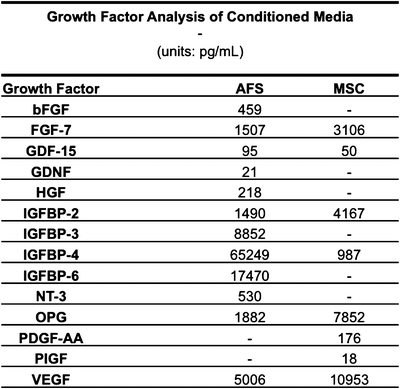
Fig. 2.5
Quantification of growth factors secreted by AFS cells and MSCs in vitro. AFS cells secrete more growth factors than MSCs. Presence of growth factor concentrations in AFS- and MSC-conditioned media were determined by proteomic arrays. Dashes indicate that growth factors were undetectable
The transient nature of this cell therapy combined with the experiments described above certainly argue strongly that the primary mechanism by which AFS cells aid wound healing biology is through secretion of trophic factors. In particular, we demonstrated the ability of AFS cells to recruit endothelial cells, behavior that was conserved in vivo in that a significantly increased number of blood vessels formed after AFS cell treatment. Indeed, other groups have found that AFS cells themselves and factors from conditioned media derived from AFS cells can induce neovascularization in ischemic skin flap models, restoring viability to the skin [37–39].
Other recruitment mechanisms may be at play as well, as suggested by the high levels of IGFBPs observed in AFS-conditioned media. Members of the IGFBP superfamily have been shown to modulate survival, migration, and proliferation of cells in the dermis and epidermis [40]. In fact, IGFBP-7 was believed to be responsible for increased migration of both fibroblast and keratinocytes in in vitro scratch assays [41]. In a separate study, IGFBP was shown to promote high levels of survival and proliferation of both HaCaT (immortalized keratinocytes) and primary human keratinocytes [42].
6 Maximizing the Effectiveness of AFS Cells in Wound Healing
The potential for AFS cells to be effective as a wound healing treatment is evident from the data presented above. However, the transient behavior observed limits that effectiveness, since when the AFS cells are no longer present, no additional GFs can be secreted. We have been recently exploring alternative hydrogel delivery vehicles to improve the logistics of cell delivery and extend the period of potency of the AFS cells and the cytokines they secrete in the wound.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


