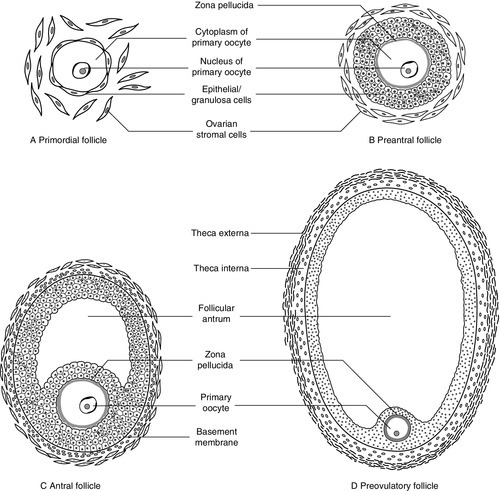
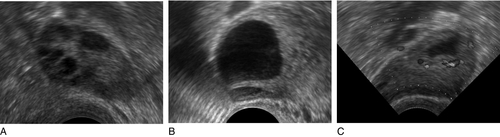
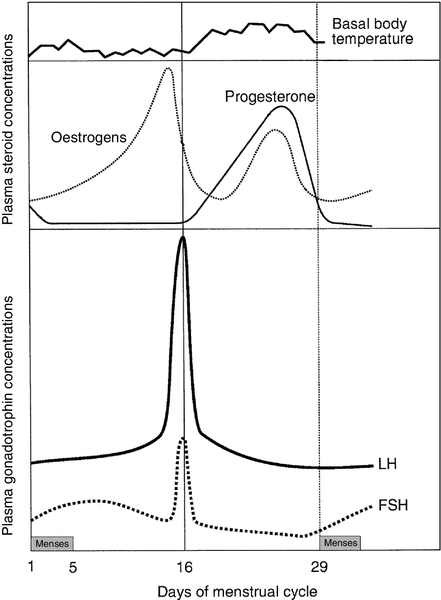
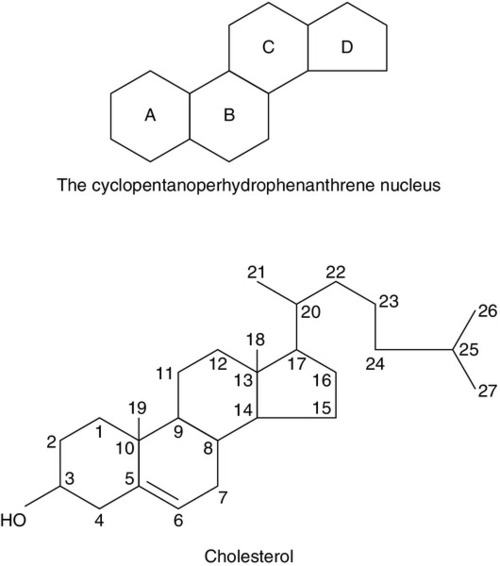
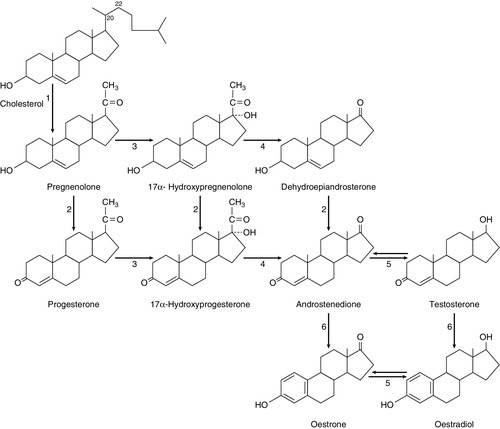
CHAPTER 22 CHAPTER OUTLINE Plasma concentrations of reproductive hormones HORMONES REGULATING REPRODUCTIVE FUNCTION Steroid secretion through the menstrual cycle Steroid hormone transport and metabolism Actions of gonadal steroid hormones Biochemical diagnosis of pregnancy Diagnosis of ectopic pregnancy Biochemical monitoring of pregnancy Screening for fetal malformation Fetal tissue sampling techniques Monitoring of maternal and fetal well-being Biochemical changes during pregnancy ORAL CONTRACEPTION AND HORMONE REPLACEMENT THERAPY Metabolic effects of oestrogens Metabolic effects of progestogens Metabolic effects of contraceptives In both normal males and females, the gonads produce steroid hormones that affect secondary sexual characteristics, the functioning of the reproductive tract and sexual behaviour. Production of gametes and hormones by the gonads is under the control of pituitary glycoprotein hormones. The hypothalamo–pituitary–gonadal axis appears to be universal throughout the vertebrates: small amounts of releasing factors from the hypothalamus elicit the release of larger amounts of glycoprotein gonadotrophins and yet larger increases in gonadal steroids. These steroids in turn influence the rate at which they themselves are produced (feedback control). The human ovaries produce female gametes and steroid sex hormones. Both functions depend largely on the monthly growth and rupture of (usually) a single ovarian follicle. At birth, some 106 immature germ cells are present in the ovaries as primary oocytes, arrested between prophase and metaphase of the first meiotic division. Each primary oocyte is surrounded by a layer of epithelial cells, the whole being known as a primordial follicle (Fig. 22.1). Primary oocytes do not complete meiosis during childhood: in fact the majority of them degenerate. FIGURE 22.1 Maturation of ovarian follicles. During childhood, the ovaries remain inactive, but at puberty, a monthly ovarian cycle is established through the interaction of the hypothalamus and pituitary with ovarian follicles and manifests itself by the onset of menstruation. At puberty, the pulsatile release of gonadotrophin releasing hormone (GnRH) from the hypothalamus stimulates the pituitary production of gonadotrophins. Growth hormone (GH) pulses released by the pituitary also increase in amplitude. This amplification of GH secretion may be regulated by the pubertal increase in the production of androgenic and oestrogenic hormones. These sex steroids stimulate skeletal growth and sexual maturation, augmenting the role of GH in promoting somatic growth and development. The human menstrual cycle is 23–39 days long. By definition, day 1 of the cycle is the first day of menstrual bleeding. The cycle is divided into follicular (or proliferative) and luteal (or secretory) phases by the event of ovulation. In cycles of different length, it is the duration of the follicular phase that varies; the length of the luteal phase is remarkably constant at 13–15 days. On day one of the cycle, several antral follicles, which are 2–9 mm in diameter, are present in both ovaries. These consist largely of a fluid-filled antrum and can be visualized by ultrasonography (Fig. 22.2A). Each antral follicle has developed from a primordial follicle by proliferation of the epithelial granulosa cells and by the appearance and coalescence of fluid-filled spaces among them (see Fig. 22.1). The earlier stages of follicle development do not require gonadotrophin stimulation, but the ‘recruitment’ of antral follicles from the preantral pool appears to be follicle stimulating hormone (FSH) and anti-Müllerian hormone (AMH) dependent. FIGURE 22.2 Vaginal ultrasonographic images of the ovary. (A) Polycystic ovary showing multiple antral type peripheral follicles; (B) Pre-ovulatory follicle 20 mm; (C) Corpus luteum with Doppler showing surrounding neovascularity. Vessels show as prominent echoes below the darker collapsed follicle. Anti-Müllerian hormone is now considered to be the main regulator of early follicular recruitment from the primordial pool. It is produced by small enlarging follicles rather than the primordial follicles themselves. Plasma concentrations of AMH do, however, appear to reflect primordial follicle numbers in humans and rodents. Through a poorly understood process of selection, one of the apparently identical antral follicles present on day 1 becomes dominant, while the others degenerate. The dominant follicle grows rapidly in the late follicular phase, reaching a maximum diameter of approximately 20 mm (Fig. 22.2B). Outside the basement membrane of the granulosa cell layer, the wall of the dominant follicle consists of the theca interna and theca externa, which have developed from ovarian stromal cells. At ovulation, the follicle collapses, releasing its fluid and the oocyte, which by now has completed the first meiotic division. The follicle subsequently refills with fluid, and blood vessels penetrate the basement membrane and vascularize the granulosa cells for the first time; further proliferation of the granulosa cells transforms what was the follicle into the corpus luteum (Fig. 22.2C). The corpus luteum has a limited life span: in the absence of conception it involutes, another dominant follicle develops and the cycle repeats itself. Insulin-like growth factor-1 has actions that regulate sex steroids through the control of the plasma concentration of sex hormone binding globulin (SHBG). Although the precise mechanism is unclear, increasing concentrations of GH are believed to exert some effects on circulating insulin concentrations. Growth hormone induces peripheral insulin resistance which leads to compensatory increases in insulin secretion and protein anabolism. Insulin regulates hepatic insulin-like growth factor 1 (IGF-1) through its effects on insulin-like binding protein 1 (IGFBP-1). Obesity is associated with increased plasma concentrations of insulin. Therefore, if excessive nutrition intake persists during childhood, it is possible that hyperinsulinaemia may lead to lower concentrations of IGFBP-1 and reduced plasma SHBG concentrations, enhancing IGF-1 and sex steroid bioavailability. Figure 22.3 illustrates the fluctuations in plasma concentrations of reproductive hormones through a typical menstrual cycle. Changes in the pulsatile release of gonadotrophins in the late luteal phase and early follicular phase appear to bring about the growth of a group of antral follicles, one of which (the dominant follicle) enlarges greatly and secretes increasing amounts of oestrogens (mainly 17β-oestradiol). Although oestrogens generally exert a negative feedback effect on gonadotrophin concentrations, high, rising oestrogen concentrations in the late follicular phase feed back positively on the hypothalamic–pituitary axis, causing a massive release of luteinizing hormone (LH) (the LH ‘surge’) and a smaller release of FSH. The LH surge triggers the resumption of meiosis and follicle rupture with oocyte release. Oestrogen concentrations fall transiently prior to ovulation. The corpus luteum secretes both oestrogens and progesterone; plasma concentrations of both hormones peak in the mid-luteal phase falling in the late luteal phase as, in the absence of conception, the corpus luteum involutes. It is likely that ovarian proteins such as inhibin and activin also play a role in regulating follicle development and steroid synthesis. These proteins and other structurally similar ‘growth factors’ may exert important paracrine effects within the ovary. FIGURE 22.3 Hormone fluctuations during the human menstrual cycle. Rising oestrogen concentrations in the follicular phase induce proliferative changes in the endometrium, causing it to thicken. Oestrogens and progesterone (from the corpus luteum) induce secretory changes in the endometrium, preparing it for pregnancy. Once deprived of hormonal support from the corpus luteum, the endometrium is shed and another cycle begins. Fertilization usually takes place in the ampullary part of the Fallopian tube, close to the ovary. Implantation occurs 6–7 days after fertilization. Human chorionic gonadotrophin (hCG) production from the conceptus acts to prolong the life of the corpus luteum so that it continues to produce progesterone, thus maintaining the endometrium. Gonadotrophin releasing hormone is a decapeptide secreted by neurons in the median eminence of the hypothalamus. It is carried in the portal hypophysial vessels from the hypothalamus to the pituitary, where it stimulates the release of LH and FSH. Concentrations of GnRH in the general circulation are very low but are assumed to be reflected by LH concentrations. The release of LH and FSH are pulsatile: the inter-pulse interval decreases from 90 to 60 min during the follicular phase until ovulation, then lengthens to once or twice a day in the late-luteal phase. Pulsatile secretion of GnRH is essential to maintain normal function, as continuous infusion causes downregulation of receptors and amenorrhoea. Administration of GnRH, modified to prolong its action, is used to induce therapeutic hypogonadism. Follicle stimulating hormone, LH and hCG are structurally similar glycoproteins. Each consists of an α- and a β-subunit, which are associated by non-covalent interactions. The α-subunits of FSH, LH and hCG (and of thyroid stimulating hormone, TSH) are identical, while the β-subunits differ, conferring hormonal specificity. The α-subunit consists of 92 amino acids. Gonadotrophins are largely metabolized in the liver; 10–15% is excreted unchanged in the urine. The β-subunit of FSH contains 118 amino acids. Carbohydrate constitutes 22–25% of the dry weight of FSH and there are four branched, mannose-rich oligosaccharide side chains covalently bound to asparagine residues, two on the α-subunit and two on the β-subunit. The biological role of the carbohydrate is partly to protect the hormone against degradation. Follicle stimulating hormone is synthesized and secreted by gonadotrophic cells of the anterior pituitary, which also synthesize LH. The primary target of FSH is the granulosa cell of the ovarian follicle. Follicle stimulating hormone stimulates differentiation of immature granulosa cells, induces cytochrome P450-dependent aromatase, induces LH receptors on granulosa cells and increases the binding of FSH by granulosa cells. The circulating half-life of FSH is ~ 4 h. Oestradiol amplifies the actions of FSH on granulosa cells. The β-subunit of LH consists of 92 amino acids. Luteinizing hormone has three carbohydrate side chains, carbohydrate accounting for approximately 15% of the dry weight of the hormone. The circulatory half-life of LH is short (20 min), contributing to the pronounced pulsatility of plasma LH concentrations. Ovarian LH receptors are found on theca cells and on mature granulosa cells. Luteinizing hormone stimulates production of androgens by the theca cells, and of oestradiol and progesterone by mature granulosa cells and corpus luteal cells. Luteinizing hormone regulates steroid biosynthesis by influencing uptake and side chain cleavage of cholesterol. The β-subunit of hCG consists of 145 amino acids. The hormone is synthesized by the syncytiotrophoblastic cells of the placenta. It has a relatively long plasma half-life (24–36 h). Human chorionic gonadotrophin interacts with the same receptors on luteal cells as does LH. It acts in the first trimester of pregnancy to maintain the corpus luteum and its ability to secrete progesterone, which supports the endometrium. After the first trimester, the placenta takes over the synthesis of progesterone from the corpus luteum. Human chorionic gonadotrophin is detectable in plasma eight days after conception and its concentration peaks around the tenth week of pregnancy. Human chorionic gonadotrophin also stimulates testosterone synthesis in the testes of male fetuses, providing stimulation for male sexual differentiation. Inhibin is a glycoprotein that has been isolated from follicular fluid in two forms comprising a common α-subunit and one of two β-subunits, βA (in inhibin A) and βB (in inhibin B). Follicular fluid also contains two dimers of the inhibin β-subunit, which are called activin: activin A is the homodimer of the βA-subunit, activin A-B is the βAβB heterodimer. Inhibin A and inhibin B have different patterns of circulation during the two phases of the ovarian cycle and play different physiological roles during follicular recruitment, maturation and ovulation. In the luteal phase, inhibin A suppresses FSH secretion. The concentration of inhibin A then decreases significantly as the corpus luteum involutes. Development of the dominant follicle is characterized by secretion of increasingly large amounts of oestradiol and inhibin A into the circulation. There is evidence that the maintenance of the follicle is affected by intraovarian signalling, with inhibins and activins acting as important paracrine messengers. During the menstrual cycle, plasma activin A concentrations vary in a biphasic manner, with the highest values occurring at mid-cycle during the luteo-follicular transition and nadirs occurring in the mid-follicular and mid-luteal phases. Prolactin is a single chain polypeptide comprising 199 amino acids. It shares a high degree of homology with growth hormone and placental lactogen. Its most potent biological form (80–90%) is monomeric; 8–20% is dimeric and 1–5% is macroprolactin. The latter is a complex of monomeric prolactin and IgG and is immunoreactive (i.e. detected in most assay systems) but biologically relatively inactive. Prolactin is synthesized by lactotroph cells of the anterior pituitary. Dopamine is the principal negative modulator of prolactin secretion; oestrogens and thyroid hormone releasing hormone increase prolactin release. Prolactin has a variety of actions in vertebrates, playing roles in processes as diverse as osmoregulation and metamorphosis. It also has a direct but variable effect on follicular development and function. Prolactin is luteotrophic in some mammals but not in man. The only definite role of prolactin in women is the postpartum initiation and maintenance of milk production. Macroprolactinaemia is a relatively common phenomenon in patients with hyperprolactinaemia in clinical practice; to ensure appropriate detection and management, all patients found to have significant hyperprolactinaemia should be screened for macroprolactinaemia (see Chapter 18). Anti-Müllerian hormone is a glycoprotein also known as Müllerian inhibitory factor (MIF) (see Chapter 21 for its role in normal and disordered sex development). It belongs to the transforming growth factor b family and is encoded on the short arm of chromosome 19. It acts through two receptors (AMHR1 and 2), which are present on the target organs in the gonads and Müllerian ducts. Measurement of AMH is used clinically for assessment of ‘ovarian reserve’ and likely response to ovarian stimulation for subfertility. It has largely superseded measurement of basal FSH and inhibin B as a marker of follicular potential, particularly as the concentrations do not change with the menstrual cycle. It can also be used as a tumour marker for granulosa cell malignancies and appears to be an excellent marker for polycystic ovarian syndrome as these patients have significantly elevated concentrations. Ovarian steroid hormones are derivatives of cholesterol and contain the cyclopentanoperhydrophenanthrene nucleus (Fig. 22.4). Oestrogens (C-18 steroids), androgens (C-19 steroids) and progestogens (C-21 steroids) contain 18, 19 and 21 carbon atoms, respectively. These compounds and the pathways involved in their biosynthesis are illustrated in Figure 22.5. FIGURE 22.4 Structural basis of steroid hormones. FIGURE 22.5 Ovarian steroid biosynthetic pathways. The numbered arrows correspond to the description of biosynthetic enzymes in the text. The numbers below refer to the enzymes and pathways labelled in Figure 22.5. 2. 3β-Hydroxysteroid dehydrogenase Δ4,5 isomerase. This catalyses both the 3β-hydroxysteroid dehydrogenation and isomerization of the double bond from ring B to ring A. Pregnenolone is thereby converted to progesterone. 3/4. 17α-Hydroxylase/17,20-desmolase. This catalyses the 17α-hydroxylation of pregnenolone and progesterone. The desmolase reaction involves the formation of a peroxide at C-20, epoxidation of the C-17 and C-20 carbons and side chain cleavage to form the C-17 oxosteroids dehydroepiandrosterone (from pregnenolone) and androstenedione (from progesterone). Both reactions are catalysed by the same enzyme. The intermediates are 17α-hydroxypregnenolone and 17α-hydroxyprogesterone, respectively. 5. 17-Oxosteroid reductase. This catalyses the conversion of a 17-oxosteroid to a 17β-hydroxysteroid and vice versa. Androstenedione and oestrone are converted to testosterone and oestradiol, respectively. 6. Aromatase. This converts C-19Δ4,3 oxosteroids to oestrogens by hydroxylation of the C-19 angular methyl group, oxidation and cleavage of the C-19 methyl group as formaldehyde, dehydrogenation of the A ring, and finally conversion of the 3-oxo group to a 3β-hydroxy group. The activity of this enzyme in granulosa cells is regulated by FSH. Oestrogens are synthesized from androstenedione, the major pathway being through oestrone. Pregnenolone is converted to androstenedione either by the Δ4 pathway (through progesterone) or by the Δ5 pathway (through dehydroepiandrosterone). The Δ4 pathway is favoured in granulosa/corpus luteal cells, while the Δ5 is favoured in theca cells. The major secreted oestrogen is 17β-oestradiol; it is in equilibrium with oestrone in the circulation. Oestrone is further metabolized to oestriol, probably mostly in the liver. Oestradiol is the most potent oestrogen of the three and oestriol the least. Before ovulation the granulosa cells are not vascularized. The theca cells produce large quantities of androgens, which diffuse into the granulosa cells and are converted to oestrogens. This concept of cooperation between cell types is called the two-cell theory. Vascularization of the granulosa cells leads to increased progesterone secretion in the luteal phase. The stromal cells of the ovaries secrete small amounts of androgens. This secretion becomes more significant after the menopause when it adds to oestrogen concentrations through peripheral conversion in adipose tissue. Most of the circulating oestrogens and progesterone is protein bound, loosely to albumin and strongly to globulins: oestrogens are carried by SHBG and progesterone by cortisol-binding globulin. Degradation of both C-18 and C-21 steroids occurs in the liver and involves hydroxylation and water solubilization by conversion to glucuronide and sulphate conjugates. Water-soluble metabolites are excreted in urine and in bile. Quantitative urinary tests were used in the past to assess concentrations of oestrogens and progestogens, but have been superseded by rapid, sensitive and specific serum immunoassays. The major excreted oestrogen and progestogen are oestradiol and pregnanediol, respectively. Oestrogens broadly maintain the functions of the reproductive tract, are responsible for the development of secondary sexual characteristics and affect sexual behaviour. Oestrogens cause myometrial hypertrophy, cause the endometrium to thicken in the follicular phase, promote secretion of large amounts of watery cervical mucus around the time of ovulation and maintain vaginal function; after bilateral oophorectomy or the menopause, the uterus and vagina shrink in size. Oestrogens produce duct growth in the breasts and are responsible for breast enlargement at puberty. Oestrogens, like androgens, increase libido in humans, apparently by a direct action on hypothalamic neurons. In general, oestrogens reduce secretion of LH and FSH (negative feedback) but, in mid-cycle, oestrogens increase LH secretion (positive feedback). In addition to their reproductive role, oestrogens have important systemic effects: they maintain bone density and skin thickness and protect against atheromatous arterial disease in premenopausal females. Synthetic oestrogen derivatives with agonist actions are used in contraceptive and hormone replacement preparations. Ethinyloestradiol is the most commonly used, particularly in contraceptives; it is potent and, unlike naturally occurring oestrogens, is active when given by mouth. Anti-oestrogen preparations are also available: clomifene citrate and tamoxifen are non-steroidal triphenylethylene derivatives with mixed oestrogen agonist and antagonist actions. Clomifene citrate is used to induce ovulation, and tamoxifen to treat oestrogen-dependent breast cancers. Progesterone causes secretory changes in endometrium already primed by oestrogen. It promotes the secretion of smaller amounts of thicker cervical mucus. It is essential for the maintenance of early pregnancy. Synthetic steroids with progesterone agonist properties are used in contraception and hormone replacement therapy. Those commonly used tend to be derivatives either of 17α-hydroxyprogesterone or of 19-nortestosterone. Both types of derivative are used in combined oral contraceptive preparations: along with ethinyloestradiol, they prevent follicle growth, promote an endometrial reaction unfavourable to implantation and render cervical mucus thick and impenetrable to sperm. Mifepristone is a derivative of norethisterone that blocks the actions of progesterone peripherally. It induces menstruation when given in the late luteal phase and induces abortion when given in pregnancy. The two main androgens in the female are testosterone and androstenedione. Dehydroepiandrosterone (DHEA) and its sulphate (DHEAS) are less important androgens. During the reproductive years, 90% of DHEA and DHEAS is synthesized by the adrenals and 10% by the ovaries. Androstenedione is derived equally from the ovaries and adrenals. Only 50% of testosterone production is glandular (roughly equal contributions from adrenals and ovaries), the remainder resulting from peripheral metabolism of weaker androgens, mostly in adipose tissue. A total of 99% of testosterone is bound and 1% is free: the great majority (78%) is bound to SHBG. This is a glycoprotein synthesized in the liver, with a carbohydrate content of 34%. It is a high-affinity, low-capacity binding protein. Plasma SHBG concentrations are raised when oestrogen concentrations are high (as in pregnancy) and in hyperthyroidism; concentrations are low in women treated with testosterone. Because SHBG-bound testosterone is relatively inert biologically, it may be useful for laboratories to measure both testosterone and SHBG, to give an indication of how much testosterone is not bound to SHBG (‘free androgen index’). Oligomenorrhoea is defined as a menstrual cycle length of greater than six weeks but less than six months. Amenorrhoea is complete absence of menstruation or cycle length greater than six months. In these conditions, ovulation does not occur or is very infrequent. Women with oligo- or amenorrhoea may seek medical assistance because their bleeding pattern is abnormal, because of infertility, because of hirsutism/virilism or with a combination of these complaints. The majority of patients with oligomenorrhoea have the polycystic ovary syndrome. The classic ‘polycystic ovary syndrome’ (PCOS) was described by Stein and Leventhal in 1935; they described an association between polycystic ovaries and oligo- and amenorrhoea in a small series of patients, most of whom were infertile or hirsute. It later became apparent that plasma concentrations of LH and/or androgens are elevated in many, but not all, women with polycystic ovaries; FSH concentrations are normal. However, it should be noted that around a quarter of women who have no gynaecological symptoms have also been found to have polycystic ovaries. Polycystic ovary syndrome is a syndrome of ovarian dysfunction. It seems most likely that changes in the pattern of gonadotrophin secretion are responsible for the failure of antral follicles to grow and for a dominant follicle to emerge and ovulate (Fig. 22.2A). It is possible to induce follicular growth in patients with PCOS by therapeutic administration of FSH. It is important to recognize that polycystic ovaries and PCOS can occasionally occur as a secondary phenomenon in patients with other endocrine disorders. These include conditions such as congenital adrenal hyperplasia and Cushing syndrome, hyperprolactinaemia and acromegaly. The revised diagnostic criteria of PCOS require any two of the following to be present: • clinical and/or biochemical evidence of hyperandrogenism • polycystic ovaries and exclusion of other aetiologies (e.g. congenital adrenal hyperplasia, androgen-secreting tumours, Cushing syndrome).
Reproductive function in the female
INTRODUCTION
PHYSIOLOGY
The ovaries
Plasma concentrations of reproductive hormones
Uterine changes
Conception
HORMONES REGULATING REPRODUCTIVE FUNCTION
Follicle stimulating hormone
Luteinizing hormone
Human chorionic gonadotrophin
Inhibin and activin
Prolactin
Anti-Müllerian hormone
REPRODUCTIVE STEROID HORMONES
Structure
Biosynthetic enzymes
Steroid secretion through the menstrual cycle
Steroid hormone transport and metabolism
Actions of gonadal steroid hormones
Oestrogens
Progestogens
Androgens
OLIGO- AND AMENORRHOEA
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


