15 UPPER DIGESTIVE SEGMENT
Upper digestive segment: Mouth, esophagus, and stomach
We have divided the discussion of the digestive system into two components or chapters: Chapter 15 focuses on the upper digestive segment and includes the mouth, esophagus, and stomach. Chapter 16 describes the lower digestive segment (small and large intestines). This division is based on the distinctive functions of the upper digestive segment (swallowing and digestion) and lower digestive segment (absorption).
MOUTH
The mouth is the entrance to the digestive tube. Ingestion, partial digestion, and lubrication of the food, or bolus, are the main functions of the mouth and its associated salivary glands. We study the salivary glands in Chapter 17, Digestive Glands.
The soft palate and uvula are lined by a nonkeratinized stratified squamous epithelium extending into the oropharynx where it becomes continuous with the pseudostratified ciliated columnar epithelium of the upper respiratory tract. The submucosa is loose and contains abundant mucous and serous glands. Skeletal muscle fibers are present in the soft palate and uvula.
Tongue
The dorsal surface of the tongue is covered by a nonkeratinized stratified squamous epithelium supported by a lamina propria associated with the muscle core of the tongue. Serous and mucous glands extend across the lamina propria and the muscle. Their ducts open into the crypts and furrows of the lingual tonsils and circumvallate papillae, respectively.
The dorsal surface of the tongue contains numerous mucosal projections called lingual papillae (Figure 15-1). Each lingual papilla is formed by a highly vascular connective tissue core and a covering layer of stratified squamous epithelium. According to their shape, lingual papillae can be divided into four types: (1) filiform papillae (narrow conical), the most abundant; (2) fungiform papillae (mushroom-shaped); (3) circumvallate papillae; and (4) foliate papillae (leaf-shaped), rudimentary in humans but well developed in rabbits and monkeys.
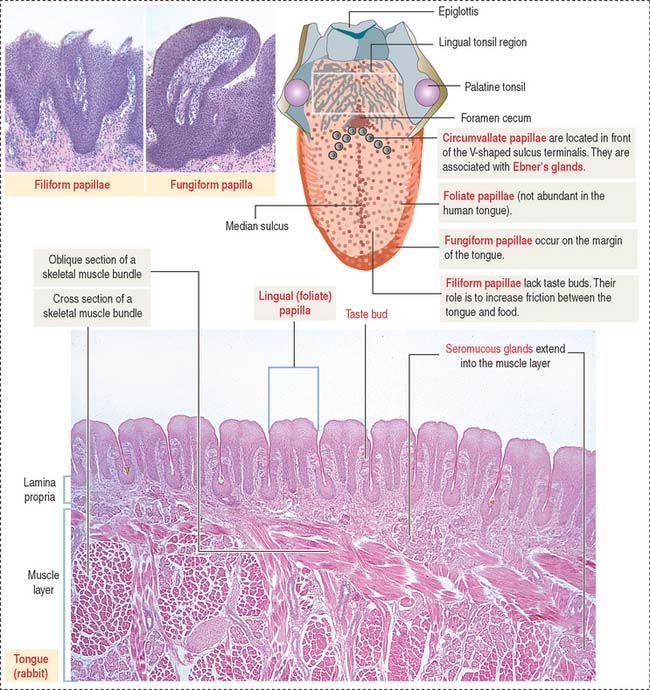
The sides of the circumvallate papilla and the facing wall of furrow contain several taste buds. Each taste bud, depending on the species, consists of 50 to 150 cells, with its narrow apical ends extending into a taste pore. A taste bud has three cell components (Figure 15-2): (1) taste receptor cells, (2) supporting cells (or immature taste cells), and (3) precursor cells (or basal cells).
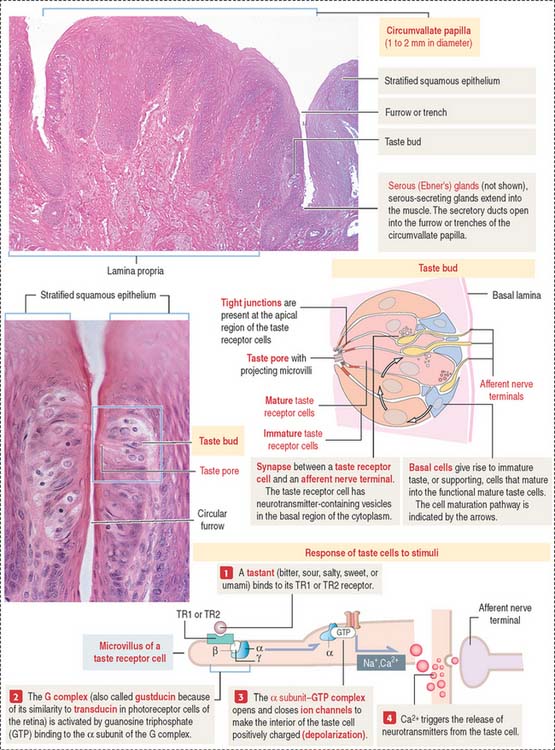
Taste is initiated when soluble chemicals, called tastants, diffuse through the taste pore and interact with the G-protein α, β, and γ subunits (called gustducin) linked to the taste receptors (designated TR1 and TR2), present in the apical microvilli of the taste receptor cells. As we discussed in Chapter 3, Cell Signaling, guanosine triphosphate (GTP) binding to the α subunit of the G-protein complex activates target molecules (ion channels in the taste receptor cells). Ionic changes within taste cells cause either depolarization (see Figure 15-2) or hyperpolarization of the receptor cells. An increase in intracellular Ca2+ triggers the release of neurotransmitters at the afferent synapse with the afferent nerve terminal. Some taste receptor cells respond to only one of the basic taste substances. Others are sensitive to more than one taste substance.
TOOTH
Each of the several types of teeth has a distinctive shape and function: incisors are specialized for cutting; canines, for puncturing and holding; and molars, for crushing.
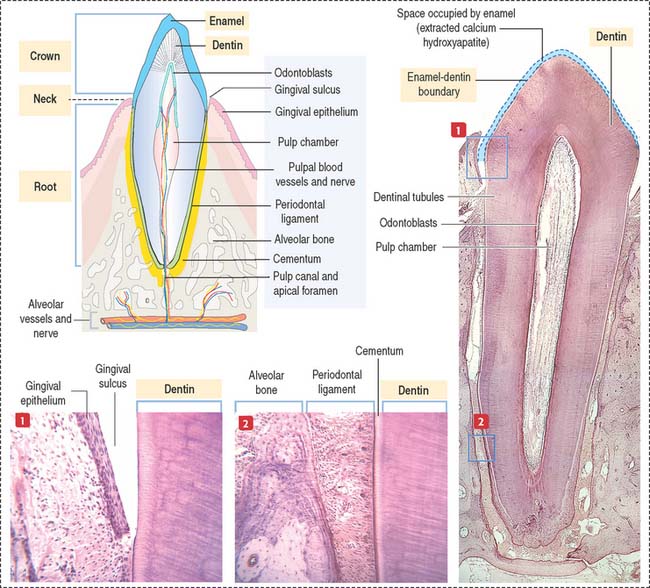
Each tooth consists of a crown and either single or multiple roots (Figure 15-3). The crown is covered by highly calcified layers of enamel and dentin. The outer surface of the root is covered by another calcified tissue called cementum. The dentin forms the bulk of the tooth and contains a central chamber filled with soft tissue, the pulp. The pulp chamber opens at the apical foramen into the bony alveolar process by the root canal. Blood vessels, nerves, and lymphatics enter and leave the pulp chamber through the apical foramen. Myelinated nerve fibers run along with the blood vessels.
TOOTH DEVELOPMENT
The ectoderm, cranial neural crest, and mesenchyme contribute to the development of the tooth (Figure 15-4). Ameloblasts derive from the ectoderm. Odontoblasts derive from the cranial neural crest. Cementocytes derive from the mesenchyme.
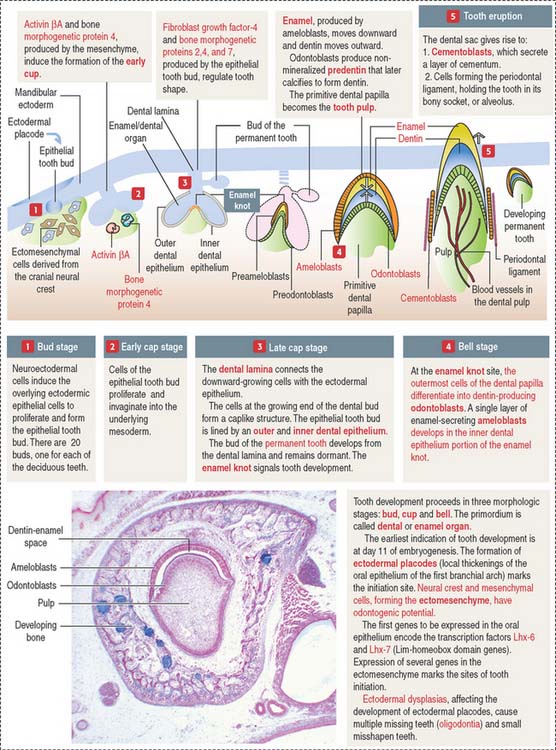
Secreted signaling molecules—activin βA, fibroblast growth factor, and bone morphogenetic proteins—mediate the interaction between the dental epithelium and the mesenchyme during tooth morphogenesis. Figure 15-4 illustrates the relevant steps of tooth development.
ODONTOBLASTS
The odontoblast is a columnar epithelial-like cell located at the inner side of the dentin, in the pulp cavity (Figure 15-5). The apical cell domain is embedded in predentin, a nonmineralized layer of dentin-like material. The apical domain projects a main apical cell process that becomes enclosed within a canalicular system just above the junctional complexes linking adjacent odontoblasts.
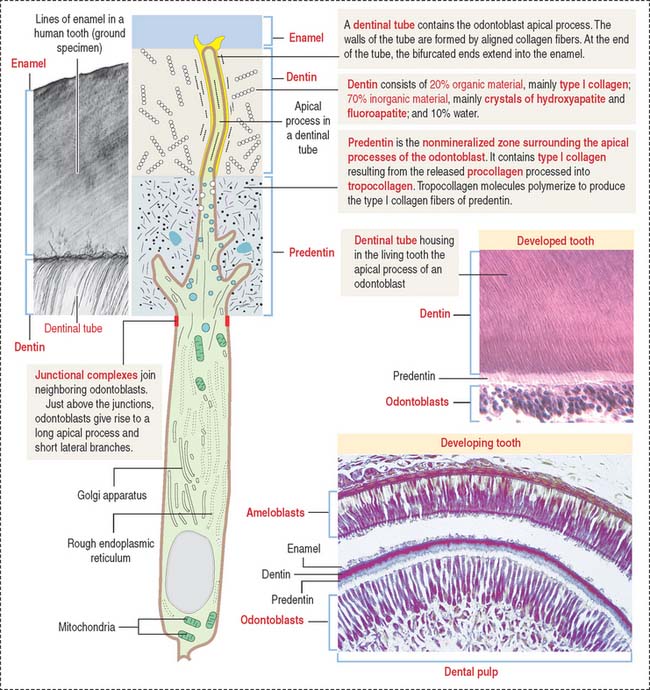
Coronal dentin dysplasia (also known as dentin dysplasia, type II) is a rare inherited autosomal defect characterized by abnormal development of dentin, extremely short roots (rootless teeth), and obliterated pulp chambers.
Cementum
The cementum meets the enamel at the cementoenamel junction and separates the crown from the root at the neck region of the tooth. The outermost layer of the cementum is uncalcified and is produced by cementoblasts in contact with the periodontal ligament, a collagen- and fibroblast-rich and vascularized suspensory ligament holding the tooth in the sockets of the alveolar bone (see Figure 15-3). The strength of the periodontal ligament fibers gives teeth mobility and strong bone attachment, both useful in orthodontic treatment.
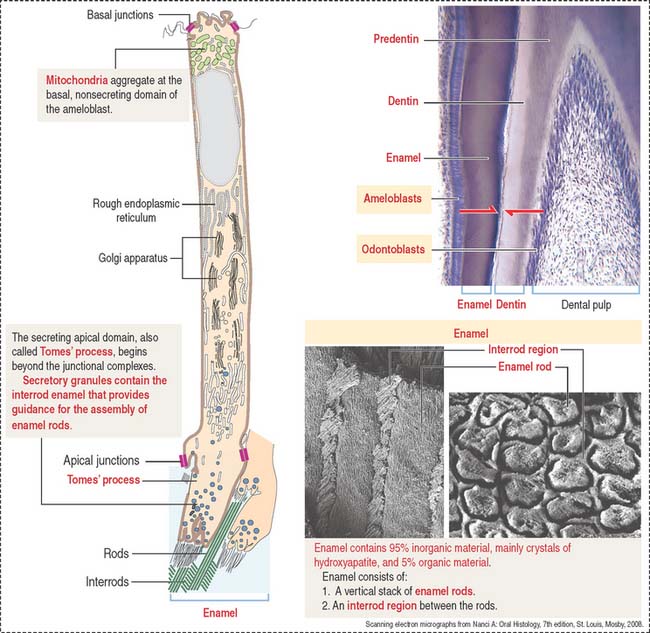
AMELOBLASTS
Ameloblasts are enamel-producing cells present only during tooth development. The ameloblast (Figure 15-6) is a polarized columnar cell with mitochondria and a nucleus present in the basal region of the cell. The supranuclear region contains numerous cisternae of rough endoplasmic reticulum and Golgi apparatus.
GENERAL ORGANIZATION OF THE DIGESTIVE, OR ALIMENTARY, TUBE
After the oral cavity, the digestive tube is differentiated into four major organs: esophagus, stomach, small intestine, and large intestine. Each of these organs is made up of four concentric layers (Figure 15-7): (1) the mucosa, (2) the sub-mucosa, (3) the muscularis, and (4) the adventitia, or serosa.
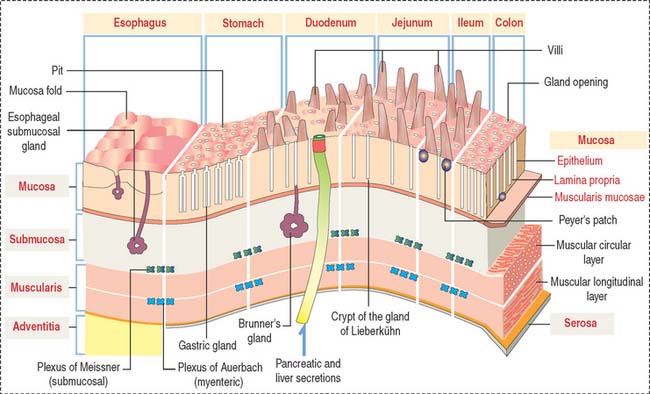
Lymphatic nodules and scattered immunocompetent cells (lymphocytes, plasma cells, and macrophages) are present in the lamina propria. The lamina propria of the small and large intestines is a relevant site of immune responses (see Chapter 16, Lower Digestive Segment).
The muscularis contains two layers of smooth muscle: the smooth muscle fibers of the inner layer are arranged around the tube lumen (circular layer); fibers of the outer layer are disposed along the tube (longitudinal layer). Contraction of the smooth fibers of the circular layer reduces the lumen; contraction of the fibers of the longitudinal layer shortens the tube. Skeletal muscle fibers are present in the upper esophagus and the anal sphincter.
Microvasculature of the digestive tube
We start our discussion with the microvasculature of the stomach. The microcirculation of the small intestine and differences from the gastric microcirculation are discussed in Chapter 16, Lower Digestive Segment (see Figure 16-3).
Blood and lymphatic vessels and nerves reach the walls of the digestive tube through the supporting mesentery or the surrounding tissues. After entering the walls of the stomach, arteries organize three arterial networks: the subserosal, intramuscular, and submucosal plexuses (Figure 15-8). Some branches from the plexuses run longitudinally in the muscularis and submucosa; other branches extend perpendicularly into the mucosa and muscularis.
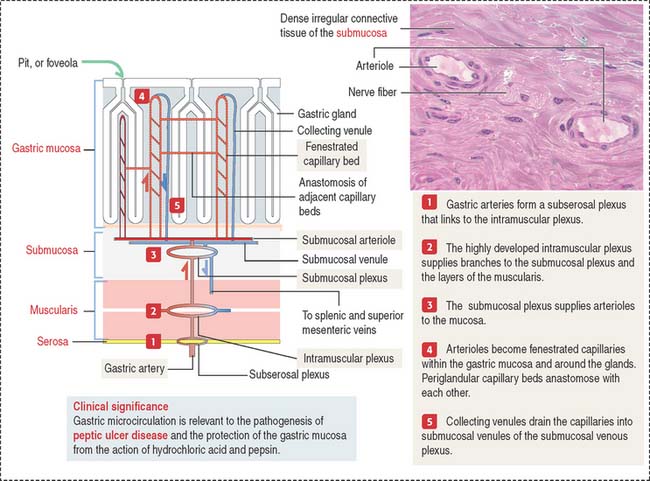
In the mucosa, arterioles derived from the submucosal plexus supply a bed of fenestrated capillaries around the gastric glands and anastomose laterally with each other. The fenestrated nature of the capillaries facilitates bicarbonate delivery to protect the surface epithelial cells against hydrochloric acid damage (see Figure 15-17).
Collecting venules descend from the mucosa into the submucosa as veins, leave the digestive tube through the mesentery, and drain into the splenic and superior mesenteric veins. Mesenteric veins drain into the portal vein, leading to the liver (see Chapter 17, Digestive Glands).



