RENAL FUNCTION: FILTRATION, SECRETION, & REABSORPTION
Renal Corpuscles & Blood Filtration
Distal Convoluted Tubule & Juxtaglomerular Apparatus
The urinary system consists of the paired kidneys and ureters, the bladder, and the urethra. This system’s primary role is to ensure optimal properties of the blood, which the kidneys continuously monitor. This general role of the kidneys involves a complex combination of renal functions:
 Regulation of the balance between water and electrolytes (inorganic ions) and the acid-base balance
Regulation of the balance between water and electrolytes (inorganic ions) and the acid-base balance
 Excretion of metabolic wastes along with excess water and electrolytes in urine
Excretion of metabolic wastes along with excess water and electrolytes in urine
 Excretion of many bioactive substances, including many drugs
Excretion of many bioactive substances, including many drugs
 Regulation of arterial blood pressure by secretion of renin
Regulation of arterial blood pressure by secretion of renin
Renin is a protease secreted into the blood that helps regulate blood pressure by cleaving circulating angiotensinogen to angiotensin I. Other renal functions include the following:
 Secretion of erythropoietin, a glycoprotein growth factor that stimulates erythrocyte production in red marrow when the blood O2 level is low;
Secretion of erythropoietin, a glycoprotein growth factor that stimulates erythrocyte production in red marrow when the blood O2 level is low;
 Conversion of the steroid prohormone vitamin D, initially produced in the epidermis, to the active form (1, 25-dihydroxyvitamin D3 or calcitriol); and
Conversion of the steroid prohormone vitamin D, initially produced in the epidermis, to the active form (1, 25-dihydroxyvitamin D3 or calcitriol); and
 Gluconeogenesis during starvation or periods of prolonged fasting, making glucose from amino acids to supplement this process in the liver.
Gluconeogenesis during starvation or periods of prolonged fasting, making glucose from amino acids to supplement this process in the liver.
Urine, the excretory product of the kidneys, passes through the ureters to the bladder for temporary storage and is then released to the exterior through the urethra.
KIDNEYS
Each kidney has a concave medial border, the hilum—where nerves enter, the ureter exits, and blood and lymph vessels enter and exit—and a convex lateral surface, both covered by a thin fibrous capsule (Figure 19–1). Within the hilum the upper end of the ureter expands as the renal pelvis and divides into two or three major calyces. Smaller branches, the minor calyces, arise from each major calyx. The area surrounding the renal pelvis and calyces contains adipose tissue.
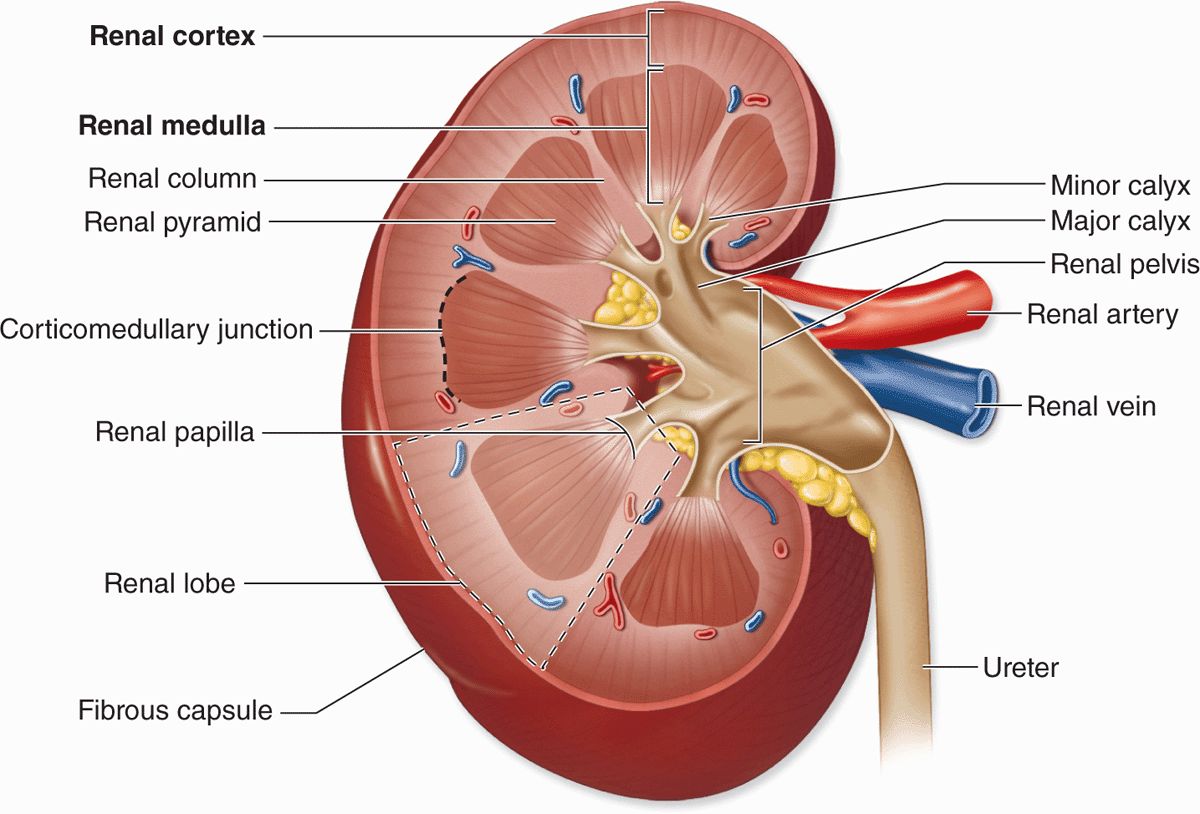
FIGURE 19–1 Kidney.
The kidney has an outer cortex, a darker region with many corpuscles and cross sections of tubules, and an inner medulla consisting of straight, aligned structures (Figure 19–1). In humans, the renal medulla consists of 8-12 conical structures called renal pyramids, which are separated by extensions from the cortex called renal columns. Each pyramid plus the cortical tissue at its base and along its sides constitutes a renal lobe. Striations extending from the medulla into the cortex are called medullary rays; these plus the attached cortical tissue are considered lobules. The tip of each pyramid, called the renal papilla, projects into a minor calyx that collects urine formed by tubules in the pyramid (Figure 19–1).
Each kidney contains around 1 million functional units called nephrons (Figure 19–2) that consist of simple, single-layered epithelium along their entire lengths. The major divisions of each nephron are the following:
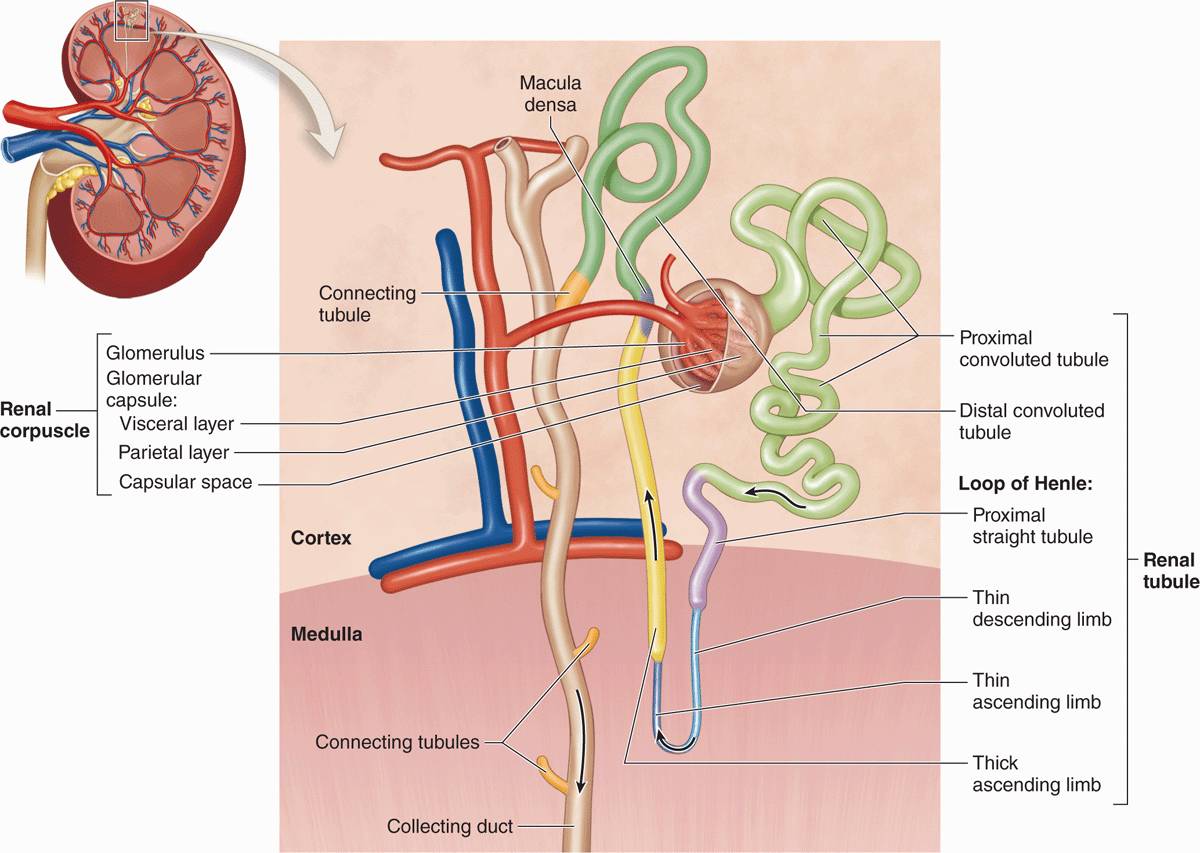
FIGURE 19–2 Nephrons.
 Renal corpuscle, an initial dilated part enclosing a tuft of capillary loops and the site of blood filtration, always located in the cortex;
Renal corpuscle, an initial dilated part enclosing a tuft of capillary loops and the site of blood filtration, always located in the cortex;
 Proximal tubule, a long convoluted part, located entirely in the cortex, with a shorter straight part that enters the medulla;
Proximal tubule, a long convoluted part, located entirely in the cortex, with a shorter straight part that enters the medulla;
 Loop of Henle (or nephron loop), in the medulla, with a thin descending and a thin ascending limb;
Loop of Henle (or nephron loop), in the medulla, with a thin descending and a thin ascending limb;
 Distal tubule, consisting of a thick straight part ascending from the loop of Henle back into the cortex and a convoluted part completely in the cortex; and
Distal tubule, consisting of a thick straight part ascending from the loop of Henle back into the cortex and a convoluted part completely in the cortex; and
 Connecting tubule, a short final part linking the nephron to collecting ducts.
Connecting tubule, a short final part linking the nephron to collecting ducts.
Connecting tubules from several nephrons merge to form collecting tubules that then merge as larger collecting ducts. These converge in the renal papilla, where they deliver urine to a minor calyx. Cortical nephrons are located almost completely in the cortex while juxtamedullary nephrons (about one-seventh of the total) lie close to the medulla and have long loops of Henle.
 MEDICAL APPLICATION
MEDICAL APPLICATION
Polycystic kidney disease is an inherited disorder in which normal cortical organization of both kidneys is lost due to the formation of multiple, large, fluid-filled cysts. The cysts may arise from any epithelial cells of the nephron and can lead to gross kidney enlargement and loss of renal function.
BLOOD CIRCULATION
As expected for an organ specialized to process the blood, the kidney vasculature is large, well-organized, and closely associated with all components of the nephron. Blood vessels of the kidneys are named according to their locations or shapes (Figure 19–3).
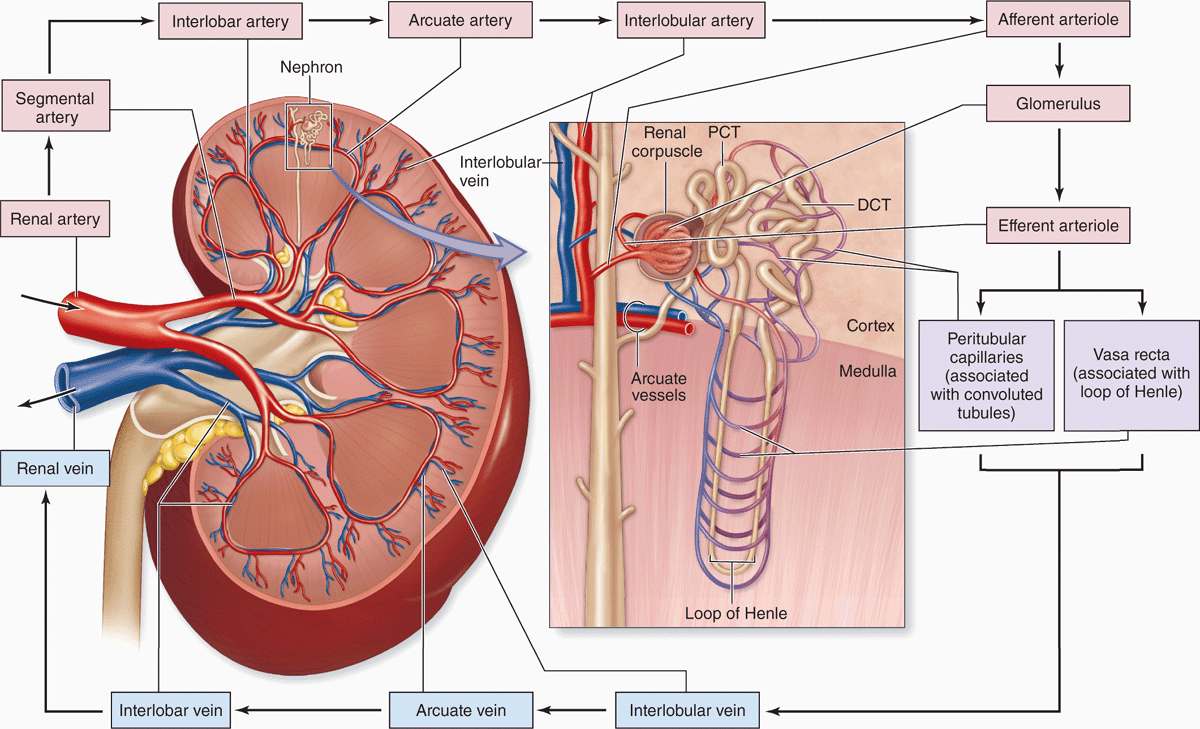
FIGURE 19–3 Blood supply to the kidneys.
Each kidney’s renal artery divides into two or more segmental arteries at the hilum. Around the renal pelvis, these branch further as the interlobar arteries, which extend between the renal pyramids toward the corticomedullary junction (Figure 19–3). Here the interlobar arteries divide again to form the arcuate arteries that run in an arc along this junction at the base of each renal pyramid. Smaller interlobular arteries (or cortical radial arteries) radiate from the arcuate arteries, extending deeply into the cortex.
From the interlobular arteries arise the microvascular afferent arterioles, which divide to form a plexus of capillary loops called the glomerulus, each of which is located within a renal corpuscle where the blood is filtered (Figures 19–3 and 19–4). Blood leaves the glomerular capillaries, not via venules, but via efferent arterioles, which at once branch again to form another capillary network, usually the peritubular capillaries profusely distributed throughout the cortex. From the juxtaglomerular corpuscles near the medulla, efferent arterioles do not form peritubular capillaries, but instead branch repeatedly to form parallel tassel-like bundles of capillary loops called the vasa recta (L. recta, straight) that penetrate deep into the medulla in association with the loops of Henle and collecting ducts. Collectively, the cortex receives over 10 times more blood than the medulla.
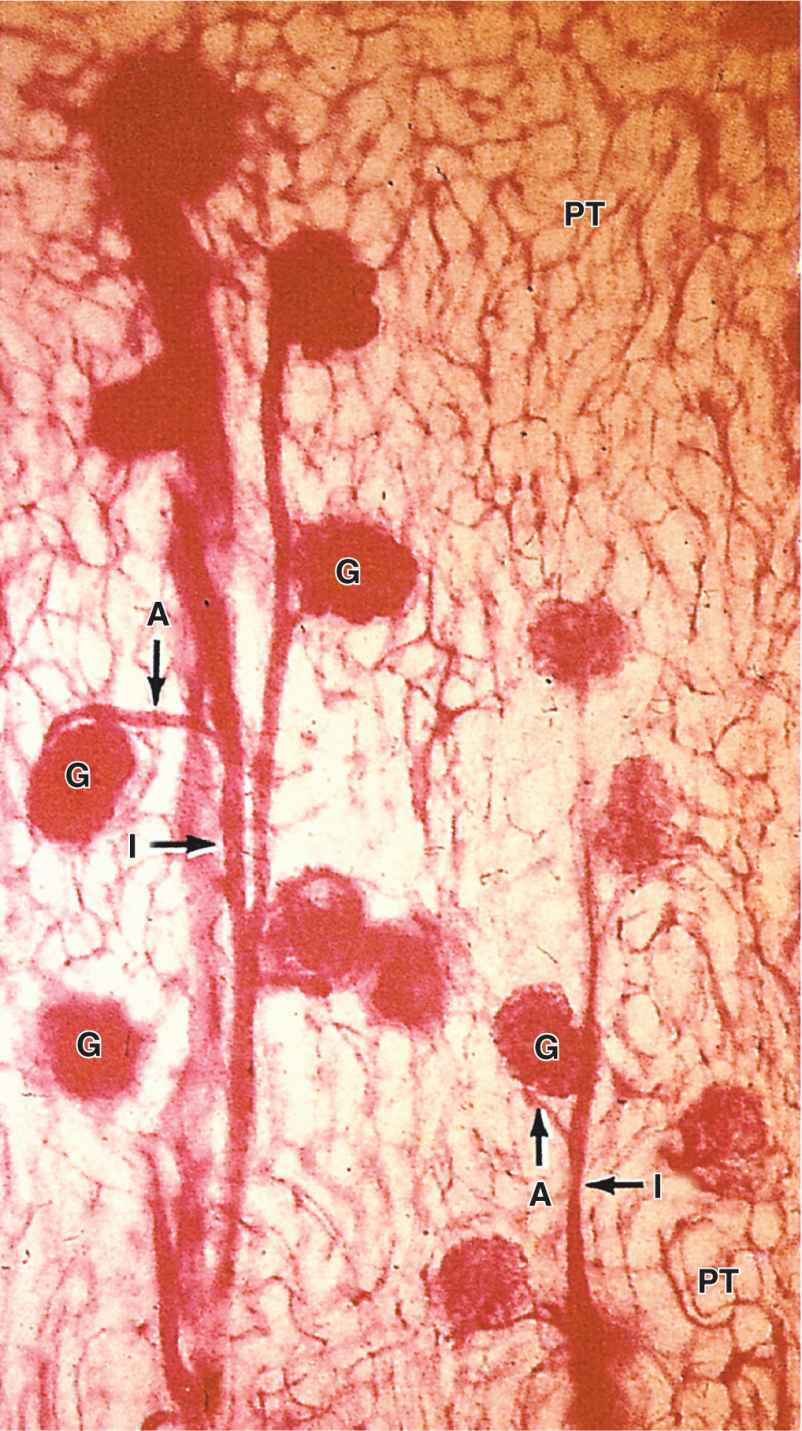
FIGURE 19–4 Microvasculature of the renal cortex.
Blood leaves the kidney in veins that follow the same courses as arteries and have the same names (Figure 19–3). The outermost peritubular capillaries and capillaries in the kidney capsule converge into small stellate veins that empty into the interlobular veins.
 MEDICAL APPLICATION
MEDICAL APPLICATIONThere are many different glomerular diseases involving the renal corpuscles, with different causes calling for different treatments. Accurate diagnoses of such disorders by pathologists require sampling of the cortex and may involve examination of the renal corpuscles by immunofluorescence light microscopy or even by TEM.
RENAL FUNCTION: FILTRATION, SECRETION, & REABSORPTION
All the major functions of the kidneys—the removal of metabolic wastes and excess water and electrolytes from blood—are performed by various specialized epithelial cells of the nephrons and collecting systems. Renal function involves specific activities:
 Filtration, by which water and solutes in the blood leave the vascular space and enter the lumen of the nephron;
Filtration, by which water and solutes in the blood leave the vascular space and enter the lumen of the nephron;
 Tubular secretion, by which substances move from epithelial cells of the tubules into the lumens, usually after uptake from the surrounding interstitium and capillaries; and
Tubular secretion, by which substances move from epithelial cells of the tubules into the lumens, usually after uptake from the surrounding interstitium and capillaries; and
 Tubular reabsorption, by which substances move from the tubular lumen across the epithelium into the interstitium and surrounding capillaries.
Tubular reabsorption, by which substances move from the tubular lumen across the epithelium into the interstitium and surrounding capillaries.
Along the length of the nephron tubule and collecting system, the filtrate receives various secreted molecules while others are reabsorbed and then enters the minor calyces as urine and undergoes excretion.
The number of nephrons decreases slightly in older adults, a process accelerated by high blood pressure. If a kidney is donated for transplant (unilateral nephrectomy), the remaining kidney undergoes compensatory growth, with cellular hypertrophy in the proximal parts of the nephron tubules and an increase in the rate of filtration, which allow normal renal function to continue.
 MEDICAL APPLICATION
MEDICAL APPLICATION
Inflammation within the glomeruli, or glomerulonephritis, which can range from acute or chronic, usually stems from humoral immune reactions. Varieties of this condition involve the deposition of circulating antibody-antigen complexes within glomeruli or circulating antibodies binding to either glomerular antigens or extraneous antigens deposited in the glomeruli. Regardless of the source the accumulating immune complexes can then elicit a local inflammatory response.
Renal Corpuscles & Blood Filtration
At the beginning of each nephron is a renal corpuscle, about 200 μm in diameter and containing a tuft of glomerular capillaries, surrounded by a double-walled epithelial capsule called the glomerular (Bowman) capsule (Figures 19–2 and 19–5). The internal or visceral layer of this capsule closely envelops the glomerular capillaries, which are finely fenestrated. The outer parietal layer forms the surface of the capsule. Between the two capsular layers is the capsular (or urinary) space, which receives the fluid filtered through the capillary wall and visceral layer. Each renal corpuscle has a vascular pole, where the afferent arteriole enters and the efferent arteriole leaves, and a tubular pole, where the proximal convoluted tubule (PCT) begins (Figure 19–5).
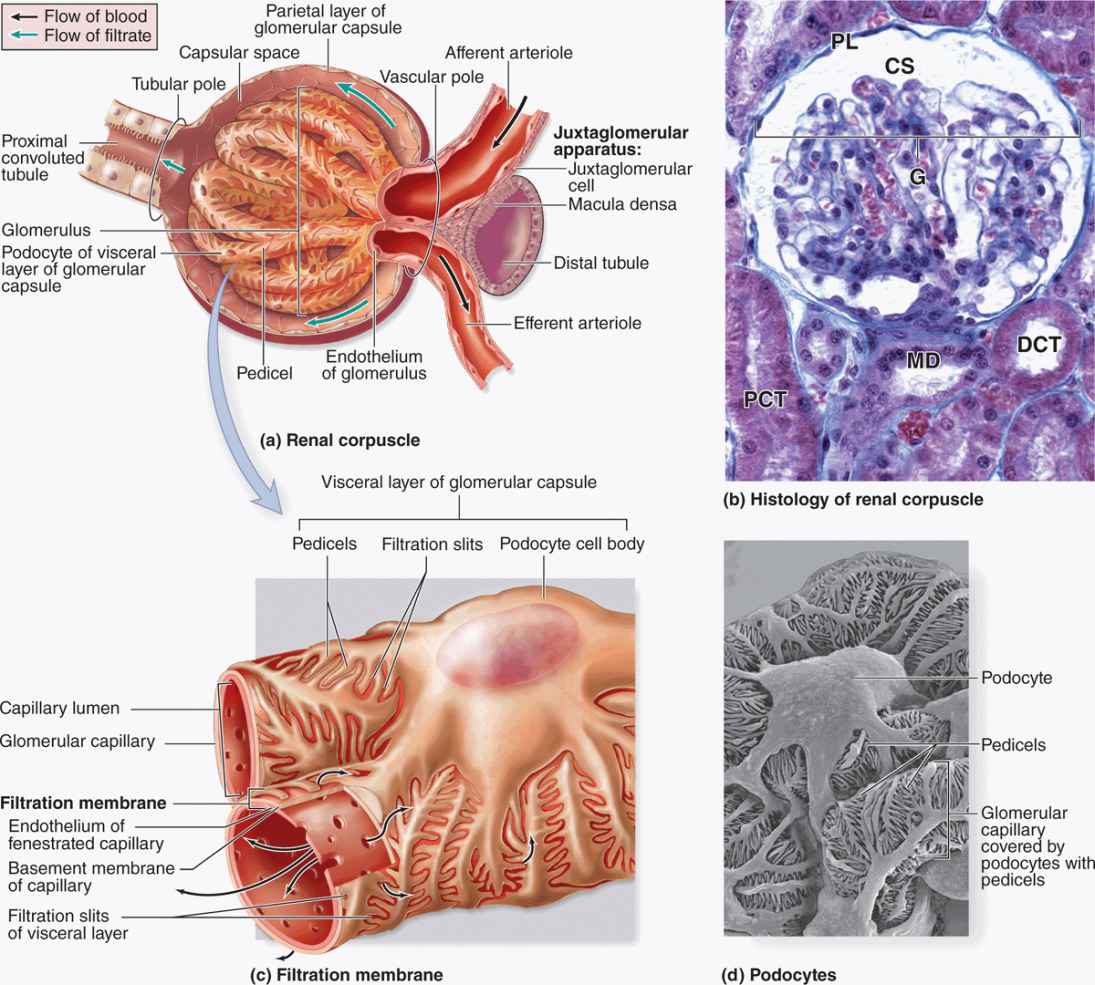
FIGURE 19–5 Renal corpuscles.
The outer parietal layer of a glomerular capsule consists of a simple squamous epithelium supported externally by a basal lamina. At the tubular pole, this epithelium changes to the simple cuboidal epithelium that continues and forms the proximal tubule (Figure 19–5).
The visceral layer of a renal corpuscle consists of unusual stellate epithelial cells called podocytes (Figures 19–5c, d), which together with the capillary endothelial cells compose the apparatus for renal filtration. From the cell body of each podocyte several primary processes extend and curve around a length of glomerular capillary. Each primary process gives rise to many parallel, interdigitating secondary processes or pedicels (L. pedicellus, little foot; Figures 19–5c, d). The pedicels cover much of the capillary surface, in direct contact with the basal lamina (Figures 19–5c and 19–6.
Between the interdigitating pedicels are elongated spaces, or filtration slit pores, 25 to 30 nm wide (Figures 19–5c and 19–6. Spanning adjacent pedicels and bridging the slit pores are zipper-like slit diaphragms (Figure 19–6). Slit diaphragms are modified and specialized occluding or tight junctions composed of nephrins, other proteins, glycoproteins, and proteoglycans important for renal function. Projecting from the cell membrane on each side of the filtration slit, these polyanionic glycoproteins and proteoglycans interact to form a series of openings within the slit diaphragm, with a surface that is negatively charged.
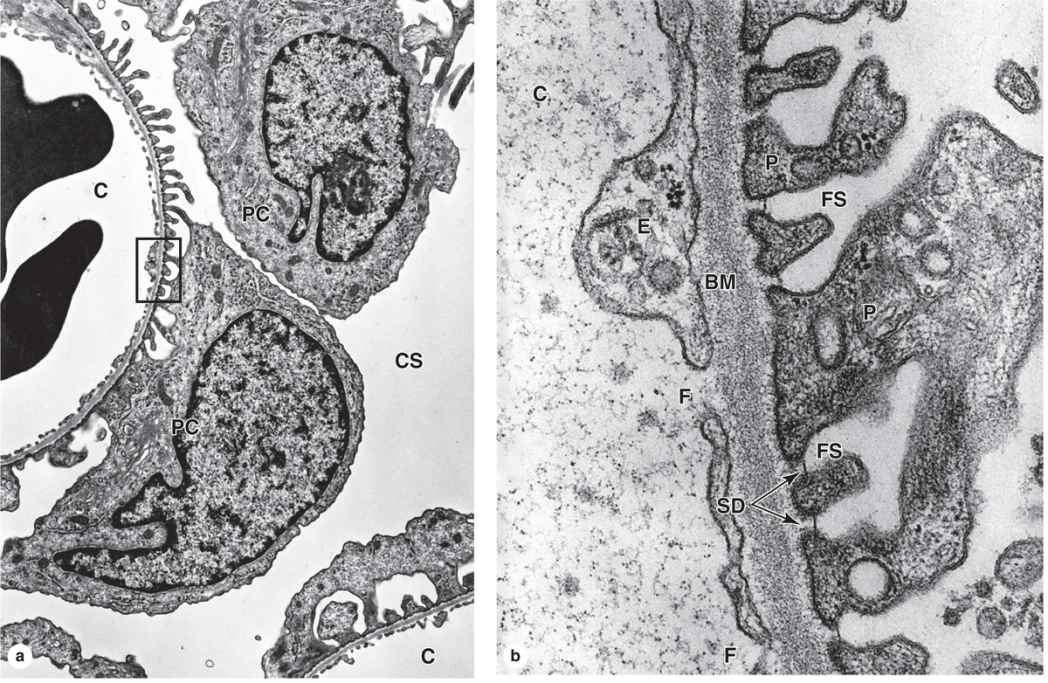
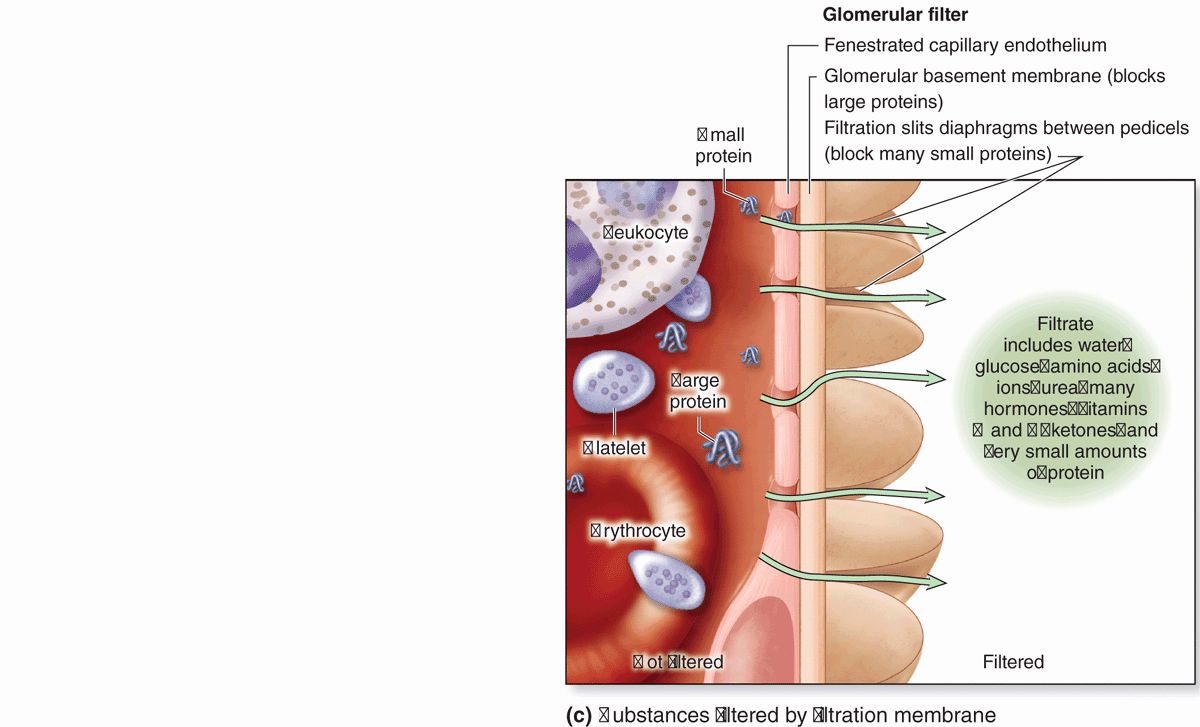
FIGURE 19–6 Glomerular filtration barrier.
Between the highly fenestrated endothelial cells of the capillaries and the covering podocytes is the thick (300-360 nm) glomerular basement membrane (GBM) (Figure 19–6). This membrane is the most substantial part of the filtration barrier that separates the blood from the capsular space and forms by fusion of the capillary- and podocyte-produced basal laminae. Laminin and fibronectin in this fused basement membrane bind integrins of both the podocyte and endothelial cell membranes, and the meshwork of cross-linked type IV collagen and large proteoglycans restricts passage of proteins larger than about 70 kDa. Smaller proteins that are filtered from plasma are degraded, and the amino acids reabsorbed in the proximal tubule. Polyanionic GAGs in the glomerular membrane are abundant and their negative charges, like those of the slit diaphragms, tend to restrict filtration of organic anions.
Filtration, therefore, occurs through a structure with three parts:
 The fenestrations of the capillary endothelium, which blocks blood cells and platelets;
The fenestrations of the capillary endothelium, which blocks blood cells and platelets;
 The thick, combined basal laminae, or GBM, which restricts large proteins and some organic anions; and
The thick, combined basal laminae, or GBM, which restricts large proteins and some organic anions; and
 The filtration slit diaphragms between pedicels, which restrict some small proteins and organic anions.
The filtration slit diaphragms between pedicels, which restrict some small proteins and organic anions.
Normally about 20% of the blood plasma entering a glomerulus is filtered into the capsular space. The initial glomerular filtrate has a chemical composition similar to that of plasma except that it contains very little protein. The glomerular filter blocks filtration of most plasma proteins, but smaller proteins, including most polypeptide hormones, are removed into the filtrate.



