● Physiologic Concepts
ANATOMY
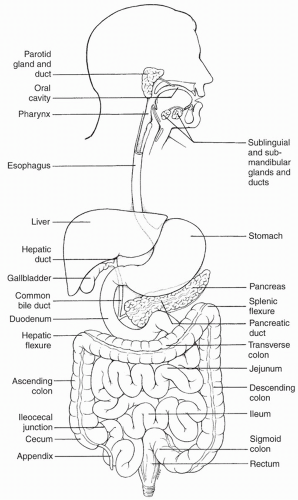
As shown in
Figure 15-1, the GI tract begins with the oral cavity, and continues into the esophagus and stomach. Food is stored in the stomach until it is released into the small intestine. The small intestine is divided into three sections: the duodenum, the jejunum, and the ileum. Digestion and absorption of food occur primarily in the small intestine. From the small intestine, food is passed to the large intestine, which consists of the colon and rectum. Accessory organs include the liver, pancreas, gallbladder, and appendix.
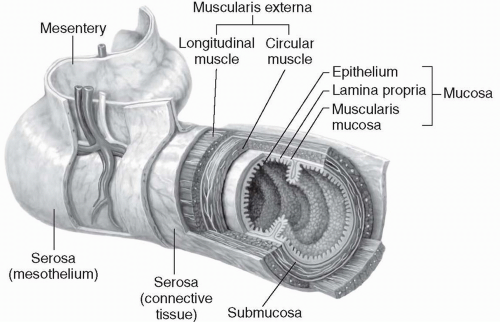
The entire GI tract is composed of several tissue layers that include:
the innermost mucosa (secreting) layer;
a submucosa connective tissue layer;
circular and longitudinal smooth muscle layers, called the muscularis externa; and
an outermost serous membrane, called the peritoneum or serosal layer, which is composed of a layer of mesothelium and a layer of connective tissue (
Fig. 15-2).
These layers are connected to one another physically and through neural connections.
Mucosa
The mucosa layer of the small intestine is the site of food absorption. The mucosa layer consists of a lining of epithelial cells, a thin connective tissue layer called the lamina propria, and an underlying layer called the muscularis mucosa. Food particles leave the gut and enter the internal environment of the body by moving across the epithelial cells.
Submucosa
The submucosa layer of the gut is a connective tissue matrix. It contains one of the two nerve networks of the gut, called the submucosal plexus, and a system of blood vessels and lymphatics. The submucosal plexus is primarily concerned with maintaining function within each very small section of the intestinal wall. For example, the submucosal plexus helps control local intestinal secretions, local absorption of foodstuffs, and local contraction of the muscularis mucosa.
Muscularis Externa
The muscularis externa contains a thick circular muscle layer, and a thinner, longitudinal muscle layer. Contraction of the circular smooth muscle causes mixing of the food in the gut. Contraction of the longitudinal layer shortens the tube. Extending all the way along the intestine, in between the circular and longitudinal smooth muscle layers, is the second neural network of the gut,
called the myenteric plexus. The myenteric plexus controls muscle contractions along the entire gut, in contrast to the localized effects of the submucosal plexus.
Peritoneum or Serosal Layer
The outer layer of the GI tract is the largest serous membrane in the body and is composed of the visceral peritoneum and the parietal peritoneum. The visceral peritoneum lines the viscera such as the intestines and stomach. The parietal peritoneum is attached to the abdominal wall. The peritoneal cavity is a fluid-filled space between the visceral peritoneum and the parietal peritoneum which facilitates movement of the abdominal structures.
NEURAL REGULATION OF THE GUT: THE ENTERIC NERVOUS SYSTEM
The two neural networks of the gut, the submucosal plexus and the myenteric plexus, make up the self-contained nervous system of the gut, referred to as the enteric nervous system. The nerves of this system fire on their own without external stimuli. Because the smooth muscle cells of the gut are connected to each other via gap junctions, firing of the nerves in one area spreads to the entire length of the gut. The neurons in the two plexuses synapse on each other, as well as on the surrounding smooth muscle cells, the exocrine glands throughout the GI tract, and the epithelial cells. They can affect contraction of the smooth muscle, the production of mucus, and the release of digestive enzymes. The neurons of the enteric nervous system include both adrenergic and cholinergic nerves, as well as nerves that release a variety of other neurotransmitters, including nitric oxide, endorphin, and various intestinal peptides. Although the firing of the enteric nervous system can occur without external input, the plexuses also receive external stimuli that influence their rate of firing.
External Neural Input
Both the myenteric and the submucosal nerve plexuses are innervated by sympathetic and parasympathetic nerves. Sympathetic fibers originate in the spinal cord between the levels of T8 and L3 and innervate the plexuses throughout the gut. They inhibit firing of the plexus, slowing the basic rhythm. Sympathetic nerves release norepinephrine in the gut. Parasympathetic nerves travel in the vagus nerve to the esophagus, the stomach, the small intestine, and the upper half of the large intestine. Other parasympathetic fibers travel in sacral divisions and innervate the distal half of the large intestine. Parasympathetic nerves release acetylcholine and stimulate firing of both the plexuses, speeding peristalsis and increasing mixing. Innervation of this distal part of the large intestine is important for stimulating defecation.
Other External Input
In addition to external neural innervation, the cells of the enteric nervous system are affected by gut hormones as well as by a variety of irritants, including those present in some foods and in certain drugs. Toxins released by infectious agents and chemicals participating in the body’s response to infection also increase the firing rate of the enteric nervous system.
THE MUSCULATURE OF THE GUT
As described previously, the GI tract is composed of an outer longitudinal layer and an inner circular layer of muscle. The longitudinal and the circular muscle layers are responsible for mixing and moving the food throughout the entire GI tract.
The longitudinal and circular smooth muscles show an inherent rate of spontaneous muscle-cell depolarization at each segment of the GI tract. These inherent depolarizations cause action potentials, resulting in muscle contractions. The contractions in each segment may vary in strength in response to internal or external nervous input, hormonal stimuli, and stretch. Although they vary in strength, gut contractions vary little in frequency. Gut contractions are slow, calcium-dependent contractions that occur over a wide range of muscle length. The contractions of the muscles at each gut segment determine the motility of that segment (i.e., the propulsion of food and secretions from one area to the next).
GUT MOTILITY
Esophageal Motility
Movement of food in the esophagus occurs by the process known as peristalsis. When food enters the esophagus, the smooth muscle is stretched; this initiates a peristaltic wave that proceeds along the length of the esophagus, propelling the food with it. When the peristaltic wave reaches the end of the esophagus, the smooth muscle at the opening into the stomach relaxes and food moves into the stomach. The end of the esophagus, called the lower esophageal sphincter (LES), is located in the abdominal cavity, below the level of the diaphragm. Because this area is not anatomically different than the rest of the esophagus, there has been a debate about whether or not the LES is a true sphincter; however, for the sake of this discussion, it will be considered a sphincter. When a peristaltic wave is not passing down the esophagus, the esophageal sphincter is relaxed and in the closed position, preventing reflux of stomach contents into the esophagus. Reflux is also prevented by the fact that the LES is in the abdominal rather than in the thoracic cavity; if this were not so, backflow of food from the high-pressure zone of the abdomen to the
low-pressure thoracic area would easily occur. By having part of the esophagus in the abdominal cavity, the pressure difference is minimized.
Stomach Motility
When food enters the stomach, the stomach also responds with a peristaltic wave. As the wave of contraction reaches the lower end of the stomach, called the antrum, the wave picks up strength, which effectively mixes the food. This wave of contraction also causes the closure of the junction between the distal end of the stomach and the beginning of the duodenum, called the pyloric sphincter. This is a true sphincter and is normally relaxed when food is not entering the stomach.
Gastric peristaltic waves occur as a result of the depolarization of the smooth muscle cells of the stomach. Pacemaker cells in the smooth muscle of the stomach depolarize continually at an inherent rate; this is called the basic electrical rhythm of the stomach. Normally, the depolarizations associated with the basic electrical rhythm are too slight to cause the muscle of the stomach to reach threshold and therefore do not lead to contractions. With increased stretch of the stomach or with neural and hormonal stimulation, the smooth muscle does depolarize to threshold and the strength of gastric peristalsis increases.
As the peristaltic waves continue in the stomach, a small amount of material is forced through the pyloric sphincter into the duodenum. The more material in the stomach, the more rapid is the rate of emptying. Eventually, all of the stomach content empties into the small intestine.
Small Intestinal Motility
Once the food, now called chyme, enters the small intestine, it continues to be mixed as a result of smooth muscle contraction. In the small intestine, the contractions result in mostly stationary mixing, with slow forward propulsion down the gut. The slow propulsion occurs as a result of segmentation. Segmentation refers to the process by which slightly more frequent contractions in the upper intestine, compared with the lower, eventually propel the chyme through the length of the small intestine. The thorough mixing during segmentation ensures that the chyme is acted upon by digestive enzymes and that it comes into repeated contact with the intestinal wall to facilitate absorption.
Large Intestine Motility
The large intestine consists of the cecum, followed by the ascending, transverse, and descending colon; the sigmoid colon; and the rectum. The appendix is a blind pouch, growing off of the cecum. The rectum ends at the anus, the exit point from the body. Contraction of the large intestine occurs at a slow rate compared with that of the small intestine. This means that food entering the large intestine takes approximately a day to travel the entire length of the
structure. A few times a day, usually after a meal, a wave of contraction, called a mass movement occurs. This is a powerful contraction that initiates the urge to defecate.
HORMONES OF THE GASTROINTESTINAL TRACT
Many GI hormones, including gastrin, secretin, cholecystokinin (CCK), glucagon-like peptide-1 (GLP-1), and glucose-dependent insulinotropic polypeptide (GIP), play important roles in the digestive function of the GI tract. Other hormones released from the stomach or intestine, including ghrelin and peptide YY (PYY), are involved in controlling appetite. These hormones and their roles are summarized in
Table 15-1.
DIGESTION OF FOOD
Digestion of food begins in the mouth with the release of saliva, continues in the stomach, and is mostly accomplished in the small intestine. The process of digestion involves enzymes that are secreted in response to specific foodstuffs and that act to break down carbohydrates into simple sugars, fats into free fatty acids and monoglycerides, and proteins into amino acids. It is only in these simple forms that these nutrients can be absorbed across the gut and used by the body.
Protein and Carbohydrate Digestion
Protein digestion begins in the stomach with the enzyme pepsin and is completed in the small intestine by the action of the pancreatic enzymes trypsin and chymotrypsin. Carbohydrate digestion begins in the mouth with the activity of the enzyme salivary amylase and is completed in the small intestine by the enzyme pancreatic amylase.
Fat Digestion
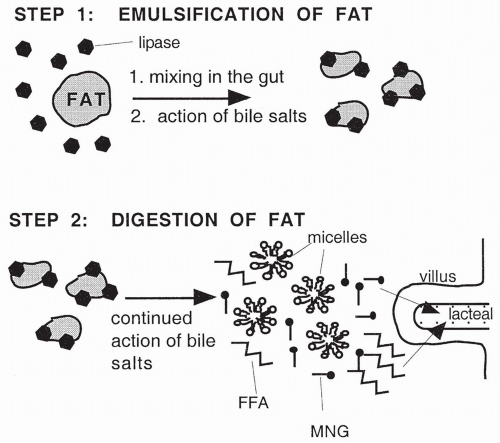
Fat digestion occurs in the small intestine, primarily as a result of the activity of the pancreatic enzyme lipase. Fats are digested by the action of lipase into free fatty acids and monoglycerides. Lipase, however, is a water-soluble enzyme; because fats are insoluble in water, their digestion by lipase would be extremely slow if it were not for emulsification, the process by which large fat complexes are broken down into smaller droplets. Emulsification increases the surface area of the fats available for digestion by pancreatic lipase. By increasing the surface area, lipase is a much more effective agent for digestion. Emulsification occurs by the mechanical mixing of the food in the intestine and by the action of bile in the small intestine. The emulsification and digestion of fat is illustrated in
Figure 15-3.
BILE
Bile is a substance produced in the liver and contains bile salts, water, cholesterol, electrolytes, and bilirubin, which is a breakdown product of hemoglobin. Although bile is continually released from the liver, it is usually stored and concentrated in the gallbladder. Bile then is released from the gallbladder and travels to the small intestine via the common bile duct, in response to the hormone CCK. In individuals who do not have a gallbladder, bile is released directly from the liver into the common bile duct in response to CCK.
Although bile contains no digestive enzymes, it does contain bile salts, the substance that serves to emulsify fats. Bile salts are phospholipids that act as detergents to break down (emulsify) fats into the small droplets. Once emulsified into droplets, lipase is then capable of digesting the fats into fatty acids and monoglycerides.
ABSORPTION OF FOOD
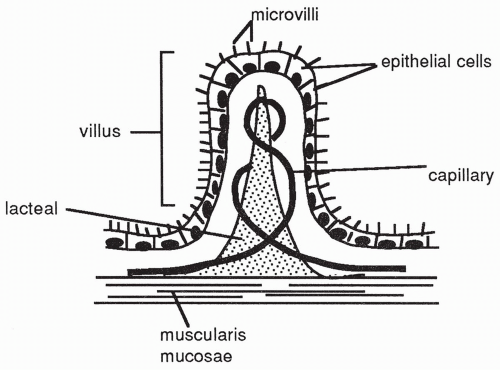
Although a small amount of lipid-soluble material may be absorbed across the stomach wall, most absorption of digested food occurs in the small intestine, across millions of fingerlike projections called villi (singular: villus). Each villus consists of epithelial cells of the mucosal layer, a capillary network, and a central lymph vessel called lacteal (
Fig. 15-4). Nerve fibers of the intrinsic plexuses and smooth muscle cells are also present. The presence of villi on the mucosal surface of the intestine increases the surface area available for the absorption of
food by at least 10-fold. Finally, each villus is topped by almost 1000 microvilli, adding even more to the enormous surface area available for the absorption of food. Certain digestive enzymes (brush border enzymes) are produced by cells of the villus as well.
Absorption of food occurs when digested food particles enter a villus from the lumen of the gut and move either into the capillary or into the lacteal, thereby gaining access to the general circulation. The movement of food particles into the epithelial cells and across the capillary or the lacteal may occur by simple or facilitated diffusion or by active transport, depending on the substance.
Absorption of Amino Acids
Amino acids and small di- or tri-peptides, the result of protein digestion, are actively transported into the epithelial cells of the villus, mostly via sodium cotransport. Once in the villus, they move into the capillary by facilitated diffusion. In the bloodstream, they are delivered to body cells, especially the muscle cells, where they are used for protein synthesis. The amino acids and small peptides not used for protein synthesis travel to the liver, where they are converted to carbohydrates or fats and are either used for energy or stored throughout the body.
Absorption of Simple Sugars
Most of the simple sugars that result from carbohydrate digestion, including glucose and galactose, are transported from the lumen of the intestine into the epithelial cells through sodium-coupled active transport. Once in the epithelial cell, these sugars then move through the basolateral membrane into the space between cells via facilitated diffusion and, from there, into the blood. Other sugars, for example fructose, move both into and out of the epithelial cells only by facilitated diffusion. Once in the blood, sugars are delivered to all body cells and are used for energy production. Sugars not used immediately for energy production can be stored as fat or glycogen in all cells, especially liver cells.
Absorption of Free Fatty Acids and Monoglycerides
Even after being digested, the absorption of free fatty acids and monoglycerides would be extremely slow if it were not for the continued action of the bile salts. Bile salts further break the emulsified fat droplets into even smaller droplets called micelles (see
Fig. 15-3). The micelles contain fatty acids and monoglycerides, bile salts and other phospholipids, cholesterol, and several fat-soluble vitamins all combined together. The micelles stay in equilibrium with a
small amount of free fatty acids and monoglycerides; these free fatty acids and monoglycerides are the substances actually absorbed into the circulation. As each molecule of free fatty acid or monoglyceride is absorbed, the micelles release replacements, thereby continuing the cycle of absorption. Without the micelles, the fat molecules would once again clump together and be unavailable for absorption.
Because the free fatty acids and monoglycerides are lipid soluble, they move by passive diffusion into the intestinal epithelial cells. In the cells, they are changed back into triglycerides, a process requiring energy. Then, triglycerides join in the epithelial cell with cholesterol and phospholipids. This complex is encased in a protein coat, exits the epithelial cell, and moves by passive diffusion into the lacteal in the center of the villi. The complex of the triglyceride, cholesterol, and phospholipid is similar to a micelle and is called a chylomicron. Chylomicrons are carried in the lymph to the thoracic duct and then enter the general circulation.
Triglycerides can be used directly as an energy source for most cells of the body, or the glycerol portion can be changed into glucose in the liver and used as an energy source. Excess triglycerides are stored in adipose tissue.
SECRETION OF MUCUS
Mucus is secreted along the entire length of the gut. Mucus is a thick substance that coats the wall of the gut and serves to protect it from being digested by the enzymes to which it is exposed. It also serves to lubricate food, allowing for easier passage. HCl, which is highly concentrated in the stomach and serves as an essential component of protein digestion, could severely compromise the gut wall integrity if mucus production were not adequate to provide protection. In addition, without the lubricating effects of mucus, stools would be hard.
RECIRCULATION OF BILE
After the bile salts deliver fatty acids and monoglycerides to the villi, some travel back into the chyme to pick up more molecules and repeat the process. Most of the remaining bile salts are eventually reabsorbed at the end of the small intestine and are recycled back to the liver via the portal vein to be used again. This process is called enterohepatic circulation.
ELIMINATION OF WASTE PRODUCTS
Absorption, primarily of water and electrolytes, continues to occur in the large intestine. Most absorption occurs in the upper half of the colon. Of the approximately 1000 mL of chyme that enters the large intestine each day, only 100 mL of fluid and virtually no electrolytes are excreted. Besides water, which makes up approximately 75% of feces, feces contain dead bacteria, some undigested fat and roughage, and a small amount of protein. Bilirubin byproducts give the feces its color.
The process of elimination, or defecation, occurs as a result of peristaltic contractions of the rectum. These contractions are produced in response to stimulation of the longitudinal and circular smooth muscles by the myenteric plexus. The myenteric plexus is stimulated by parasympathetic nerves traveling in sacral segments of the spinal cord. Mechanical stretching of the rectum with
stool is also a strong stimulator of peristalsis. When a peristaltic wave is initiated, the internal anal sphincter, a smooth muscle, relaxes. If the external anal sphincter is also relaxed, defecation occurs. The external anal sphincter is a skeletal muscle and thus under voluntary control. In fact, relaxation of the internal sphincter causes reflex contraction of the external sphincter in all individuals except babies and some people who have spinal cord transection. Reflex contraction of the external sphincter effectively stops defecation. If the defecation reflex occurs at an acceptable time, the reflex contraction of the external sphincter can be consciously reversed to allow defecation.
HUNGER AND THE INGESTION OF FOOD
Hunger is controlled by an area of the brain in the lateral hypothalamus. Stimulation of this area causes a strong desire to seek out and to eat food. The lateral hypothalamus receives numerous inputs that can stimulate hunger. For instance, hunger can be stimulated by the occurrence of hunger contractions in the stomach. These contractions appear to increase in frequency and intensity the longer the stomach remains empty. The exact mechanism by which they occur is unclear.
Hunger is also stimulated by a fall in blood nutrients, such as amino acids, fats, and glucose, and by a rise in the hormones that accompany nutrient deficit (e.g., glucagons and ghrelin). A decrease in the level of hormones present when food is plentiful may also stimulate hunger (e.g., decreased insulin and PYY). Input to the hypothalamic hunger center can include input from other areas of the brain as well. For instance, higher brain centers can stimulate hunger in response to certain situations or experiences. Likewise, input from the emotional center of the brain, the limbic system, may also stimulate hunger, as may different smells activating from the olfactory center.
Conversely, the ventromedial nucleus of the hypothalamus appears to be the site where satiety, the opposite of a hunger drive, occurs. This center is influenced by the fullness of the stomach and blood levels of nutrients and hormones, and also in the opposite direction, as is required for hunger stimulation. Emotions and habits may also influence the satiety center.
TESTS OF GASTROINTESTINAL FUNCTIONING
Barium Contrast x-Ray Films: Upper and Lower Gastrointestinal Series
In these tests, a radiopaque solution is introduced into the upper or the lower GI tract, and then x-ray films are obtained to follow its progress. This technique is able to identify the positions and sizes of the GI structures and any obstructions that are present; however, its ability to identify ulcers, fissures, or early stage cancers is poor.
Endoscopy
Endoscopy is the process whereby a thin, rigid or flexible scope is passed into the GI tract to visualize the esophagus (esophagoscopy), stomach (gastroscopy), upper small intestine (duodenoscopy), large intestine (colonoscopy), or sigmoid colon (sigmoidoscopy). With this instrument, the walls of the GI tract can be visualized, allowing identification of ulcerations, blockages, and other irregularities. Special tools at the end of the scope allow tissue to be sampled for biopsy and culture.
Whether patients should have colonoscopy or sigmoidoscopy for screening of colon cancer depends on personal risk factors, including age, family history of GI or other cancers, and personal history of polyps or cancer. With colonoscopy, the practitioner can fully visualize the entire large intestine. Because many colon cancers develop in the sigmoid colon and because sigmoidoscopy is usually accomplished without general anesthesia, this procedure may be recommended for screening in low-risk populations.
Endoscopic Retrograde Cholangiopancreatography (ERCP)
The gallbladder and pancreas are accessory organs of the GI system and play a major role in the production and storage of bile salts that are necessary for the emulsification of fat. When a problem with either of these organs is suspected, an ERCP is performed. ERCP is similar to the endoscopy but the focus is on the pancreatic and bile ducts. Dye is injected and the ducts are visualized. Stone removal, biopsy, and sphincterotomy are possible with this procedure.