9 The skin is the external body surface and provides protection against a wide variety of external threats, including mechanical, water loss, biological, ultraviolet light and chemical. The most frequent mechanical insult is the frictional and shearing forces experienced by the soles and ventral aspect of the toes in walking and, to a lesser extent the palms and ventral aspects of the fingers during use of the hands. In these areas, skin structure is adapted to resist these shearing forces (see Fig. 9.20). The skin is the largest sensory organ in the body, containing a range of different receptors for touch, pressure, pain and temperature (see Ch. 7). As a regional variation in structure, sensory receptors are most numerous in skin which has the most physical contact with solid objects in the environment, such as soles and palms, fingers and toes (see Fig. 7.24), or with specialised sexual functions. The skin has three main layers: • The epidermis is a continuously proliferating stratified squamous epithelium which produces a non-living surface layer of the protein keratin, with associated lipid which is in direct contact with the external environment and is constantly shed. • The dermis consists of fibrous and fibroadipose tissue which supports the epidermis, both physically and metabolically. It contains blood vessels, nerves and sensory receptors. • The subcutis or hypodermis or panniculus is the layer beneath the dermis and usually consists of adipose tissue with supporting fibrous bands (septa). This layer contains the larger vessels which supply and drain the dermal blood vasculature. FIG. 9.1 Skin architecture FIG. 9.2 Epidermis FIG. 9.3 Keratinocytes (illustrations (a) and (b) opposite) FIG. 9.4 Dermo-epidermal junction FIG. 9.5 Langerhans cells
Skin
Introduction
Skin Structure
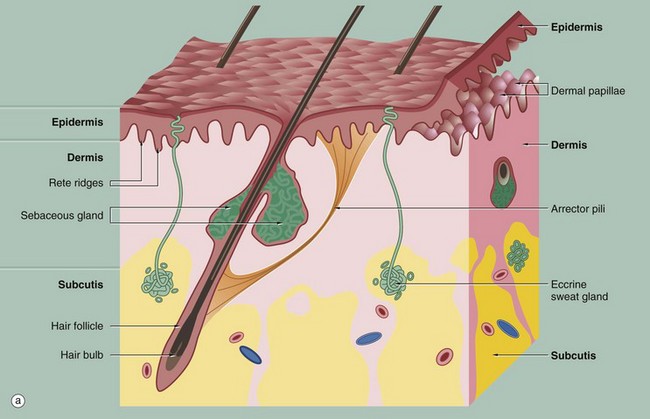
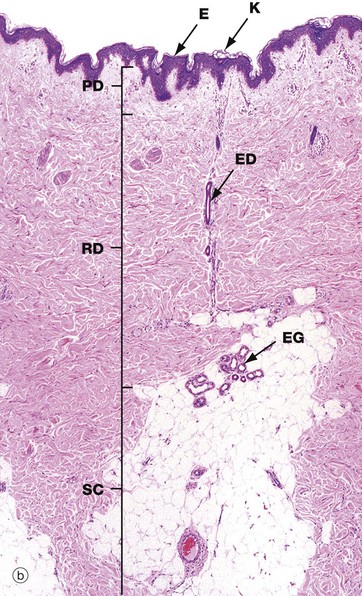
(a) Diagram (b) H&E (LP)
These illustrations show the basic structure of the skin, with the three component layers: epidermis, dermis and subcutis.
The surface layer in contact with the exterior is the epidermis E, a highly specialised self-regenerating stratified squamous epithelium which produces a non-living surface rich in a protein, keratin K, that is tough and protective and is also partially water resistant (see Ch. 5). The epidermis also contains non-epithelial cells: melanocytes produce melanin pigment to protect against UV light, Langerhans cells act as antigen-presenting cells and induce immune responses to new antigens and Merkel cells act as touch receptors. The epidermis is tightly bound to the underlying dermis by a specialised basement membrane. Additional resistance to frictional shearing force is provided by a series of epidermal downgrowths (rete ridges) which extend into the superficial dermis, with their papillary dermal mirror images projecting upwards (dermal papillae) to provide stronger tethering. These are most developed where exposure to shearing forces is almost constant (e.g. sole, palm).
The dermis immediately adjacent to the epidermis is called the papillary dermis PD; it has relatively fine collagen fibres and contains numerous small blood vessels, sensory nerve endings and sensory structures. The reticular dermis RD is the deeper tough layer of horizontally arranged collagen and elastin fibres with fibroblasts.
The deepest layer is the subcutis SC, also called the panniculus or hypodermis. It is a layer of adipose tissue often compartmentalised by fibrous septa, extending downwards from dermis to the underlying structural connective tissue fascia. The subcutis acts as a shock absorber and thermal insulator as well as a fat store.
The dermis and subcutis contain an assortment of skin adnexa (appendages) such as hair follicles, sebaceous glands, eccrine (sweat) glands EG and ducts ED and, in some areas, apocrine glands.
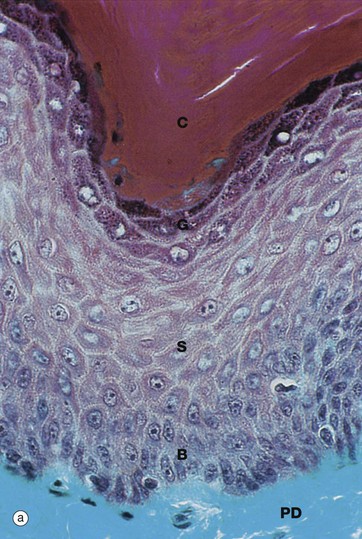
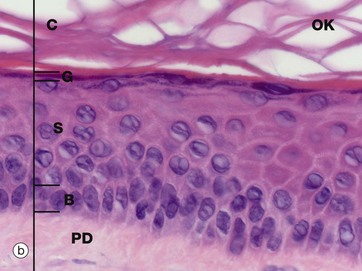
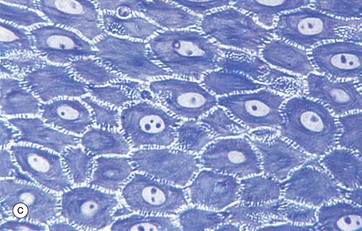
(a) Masson trichrome (HP) (b) H&E (HP) (c) Epoxy resin section, toluidine blue (HP)
Micrographs (a) and (b) show epidermis. The cells of the epidermis are called keratinocytes. The basal layer of keratinocytes (stratum basale) B proliferates continuously with repeated mitotic divisions. This provides cells for a progressive process of displacement towards the surface (upward migration), with associated maturation to renew the other layers. The basal cells are arranged as a single layer of cuboidal or low columnar cells. They are attached to the basement membrane (not seen in these preparations) on their dermal (basal) surface. This basal surface is irregular; the basal cells have a highly indented and folded basal cell membrane with numerous hemi-desmosomes.
Superficially, the basal cells are attached to and mature into the cells of the stratum spinosum S which forms the majority of the epidermis. The stratum spinosum is also known as the prickle cell layer. It is multilayered and composed of polyhedral-shaped keratinocytes with round-oval nuclei, prominent nucleoli and cytoplasm, forming a pavement-like pattern. These cells synthesise cytoplasmic intermediate filaments called cytokeratins which accumulate in aggregates called tonofibrils made up of bundles of tonofilaments. These tonofibrils bind to the numerous desmosomes that form strong contacts between adjacent keratinocytes. In appropriate preparations as in micrograph (c), the desmosome junctions are seen as prickles or spines between the cells, hence the name for this layer.
The keratinocytes mature into the stratum granulosum G or granular layer. Here they acquire dense basophilic, keratohyaline granules which contain proteins rich in sulphur-containing amino acids (cysteine) and proteins such as involucrin which interact with the cytokeratin tonofibrils in the final maturation. The combination of tonofibrils with keratohyaline granule proteins produces keratin, in a process called keratinisation. Progressing towards the surface, the cells lose their nuclei and cytoplasm, becoming flattened interconnected keratin squames (plates/flakes of keratin) which comprise the surface coating of the skin, the stratum corneum C.
These keratin squames connect at their edges, and in transverse sections form a folded basket-weave pattern called orthokeratosis OK. The squames are water repellent, in part because they are coated with lipid-containing anti-wetting agents synthesised during maturation in the granular layer.
Micrograph (a) illustrates skin from a friction-prone area (sole of foot) where keratin production is enhanced. The stratum spinosum has more layers, the granular layer G is thicker and more prominent and the stratum corneum C is thick with dense compact keratin. These changes are a non-specific response to friction.
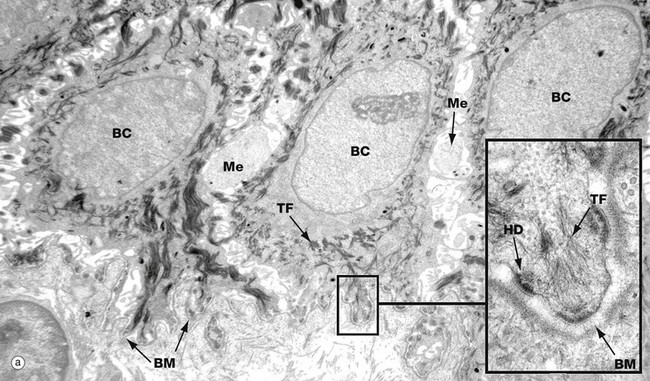
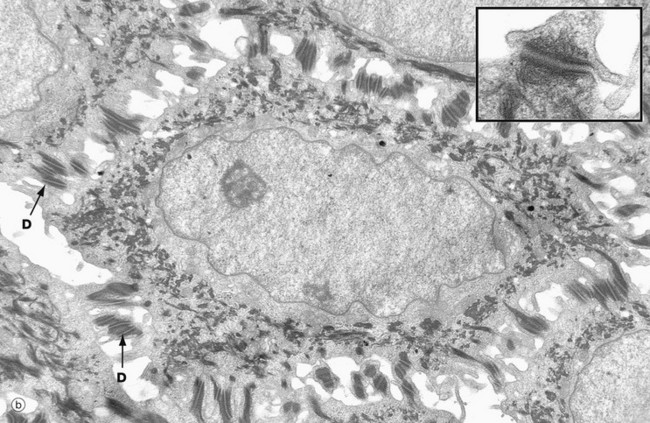
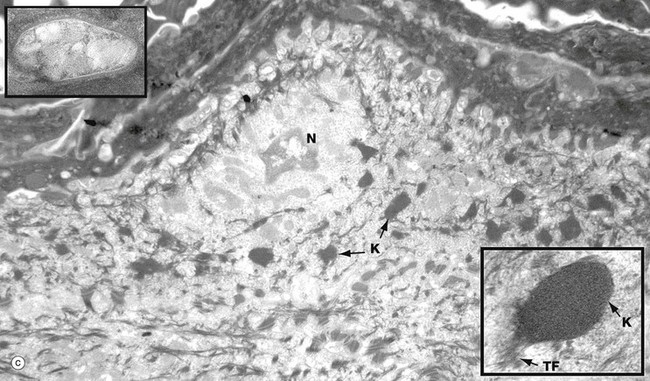
(a) Basal keratinocytes, EM ×10 000, inset ×30 000 (b) Keratinocyte from prickle cell layer, EM ×15 000, inset ×40 000 (c) Keratinocyte from granular layer, EM ×12 000, inset ×50 000
Micrograph (a) shows a row of three basal cells BC sitting on the epidermal basement membrane BM, which can only just be discerned as a convoluted line in the low-magnification main picture. The basal cells are cuboidal with prominent nuclei, nucleoli, and a perinuclear zone containing ribosomes and mitochondria. Tonofibrils TF are present towards the periphery of the cell.
The inset shows the linkage of the basal surface of the cell to the basement membrane BM by hemi-desmosomes HD into which tonofibrils TF are inserted. Basal cells link to each other and to overlying keratinocytes of the prickle cell layer by desmosomes. The pale processes in the spaces between basal cells are cytoplasmic melanocyte processes Me.
Micrograph (b) shows a single keratinocyte from the prickle cell layer. It has abundant electron-dense tonofibrils which extend into the many desmosomes D that link it firmly to adjacent keratinocytes. The inset shows a desmosome linking the cytoplasmic processes of two keratinocytes. Electron-dense tonofibrils from each cell attach to the desmosome.
Micrograph (c) shows a flattening keratinocyte in the granular layer immediately beneath the keratin layer. The cell is keratinising with an ill-defined remnant of the nucleus N. The cytoplasm contains both linear cytokeratin tonofibrils and electron-dense ovoid keratohyaline granules K, one of which is shown merging with tonofibrils TF at higher magnification in the inset bottom right. Also present but not clearly visible in the low-magnification micrograph are lamellated pale-staining ovoid bodies (about 500 nm long) called keratinosomes or Odland bodies; one is shown at higher magnification in the inset top left. They contain a hydrophobic glycolipid which when released coats and binds together the keratin flakes, rendering them relatively water repellent.
(a) Diagram (b) EM ×64 000
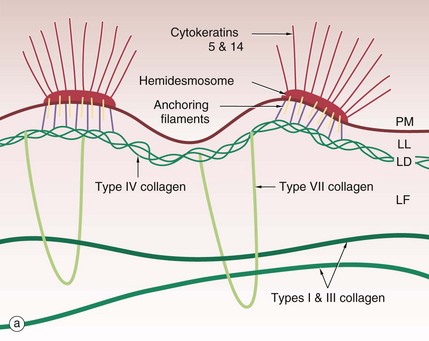
The basement membrane at the junction of epidermis and dermis, known as the dermo-epidermal junction, is a specialised structure which binds epidermis to dermis. The major components are illustrated in the diagram (a).
Cytoplasmic tonofibrils, consisting of cytokeratins 5 and 14, bind to hemidesmosomes in the plasma membrane PM at the base of the basal cells of the epidermis. The hemidesmosomes contain multiple transmembrane and binding proteins including type XVII collagen and integrins. Binding to these on the outside of the plasma membrane are the anchoring filaments, including nidogen-1 and laminins. These correspond to an electron-lucent layer on electron microscopy called the lamina lucida LL. This appears to be the most easily disrupted of the basement membrane layers.
These anchoring filament proteins bind to a type IV collagen layer, type IV collagen being a specialised network/mesh-forming basement membrane collagen. On EM this corresponds to the lamina densa LD, an electron-dense layer.
Type VII collagen is a basement membrane specialised collagen which forms loops extending from the type IV collagen layer around fibres of structural types I and III collagens in the dermis, thereby tethering the basement membrane to the dermis; these are the anchoring fibrils. These form the lamina fibroreticularis LF, a poorly defined layer on EM.
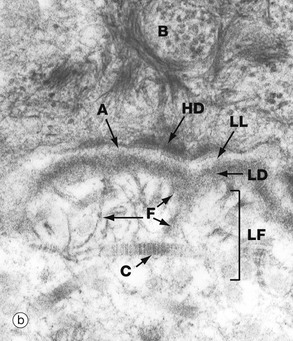
The electron micrograph (b) shows part of a basal cell B, basement membrane layers of lamina lucida LL, lamina densa LD and lamina fibroreticularis LF, in addition to fibrils of type VII collagen F which tie to the type I collagen fibres C in the dermis. There are also finer fibrillin microfilaments which link to dermal elastic fibres. Basal cell hemidesmosomes HD can be seen at the plasma membrane with their fine anchoring protein filaments A which cross the lamina lucida.
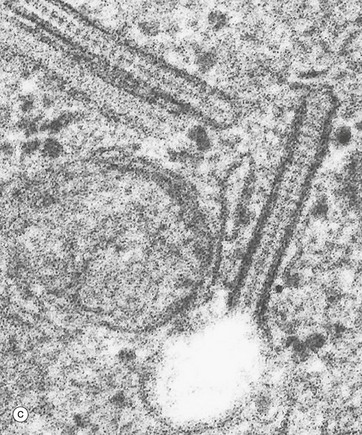
(a) H&E (HP) (b) Immunohistochemistry for CD1a (HP) (c) Birbeck granule, EM ×100 000
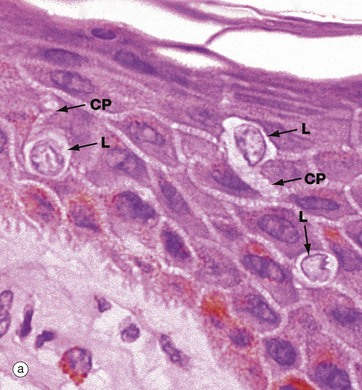
Langerhans cells are intra-epidermal antigen presenting cells, historically referred to as histiocytes. They are present in all layers of the epidermis but are most easily recognised in the prickle cell layer. They are also present in the upper dermis, particularly around small blood vessels. When stimulated, they migrate to dermis and then via lymphatics to lymph nodes.
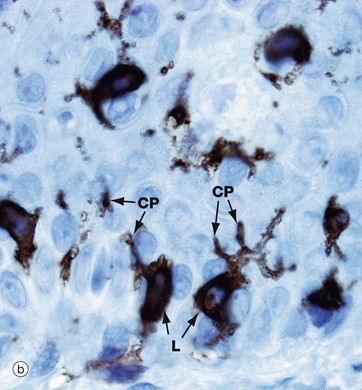
Micrograph (a) shows pale-staining Langerhans cells L in the epidermis; they have irregularly lobulated nuclei and almost clear cytoplasm. Cytoplasmic processes CP extend from the cells and insinuate between keratinocytes of all layers. The extensive network of cytoplasmic processes is highlighted in the immunohistochemical preparation shown in (b).
Electron micrograph (c) shows the characteristic cytoplasmic organelle of Langerhans cells, the Birbeck granule, a rod-like structure with regular cross-striations, one end of which frequently distends in a vesicle so that they resemble a tennis racket. They are part of the endosome system. Their exact function is not known.![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Basicmedical Key
Fastest Basicmedical Insight Engine
