CHAPTER 15 Overview of the nervous system
The human nervous system is the most complex product of biological evolution. The constantly changing patterns of activity of its billions of interactive units represent the fundamental physical basis of every aspect of human behaviour and experience. Many thousands of scientists and clinicians around the world, whether driven by intellectual curiosity or the quest for better methods of disease prevention and treatment, have studied the nervous system over many years. Despite the enormous amount of data available, our understanding of complex neural organization and function is far from complete, as is our ability to deal with its many pathologies. Nevertheless, research on the nervous system is one of the most active areas of contemporary biology and medicine and rapid advances on numerous fronts bring with them the realistic prospect of better prevention and treatment of many neurological disorders in the future.
The functional capabilities of the nervous system are a product of its vast population of intercommunicating nerve cells or neurones, estimated to number in the order of 1010 (Ch. 3). In brief, neurones encode information, conduct it, sometimes over considerable distances, and then transmit it to other neurones or to non-neural tissues such as muscles or glandular cells. Most neurones consist of a central mass of cytoplasm within a limiting cell membrane, the cell body or soma, from which extend a number of branched processes, or neurites (Fig. 15.1). One of these, the axon, or nerve fibre, is usually much longer than the others and conducts information away from the cell body. The other processes are termed dendrites and these typically conduct information towards the soma (cell body). The nerve cell membrane is polarized, the inside of the cell being around −70 mV negative with respect to the outside. Information is coded in the form of patterns of transient depolarizations and repolarizations of this membrane potential, known as action potentials or nerve impulses. These are conducted along the axon, which may have collateral branches that permit information to be distributed simultaneously to several targets. Axons possess specialized endings, or axon terminals, which come into close apposition with the membrane of the target cell at synapses, where information passes from one cell to another (Fig. 15.2). Axon terminals may form synaptic contacts with dendrites (axodendritic), cell bodies (axosomatic), other axons (axoaxonic) or non-neural tissue such as muscle cells (neuromuscular junction). Transmission of information to other cells occurs when action potentials cause the release of specific neurotransmitter substances, stored in synaptic vesicles within the presynaptic nerve terminal. The neurotransmitter binds to specialized receptors that are located on the postsynaptic target cell membrane and, depending upon the nature of the chemical and the receptor, either elicits an excitatory (depolarizing) or inhibitory (hyperpolarizing) response, or may modulate intracellular second messenger systems.
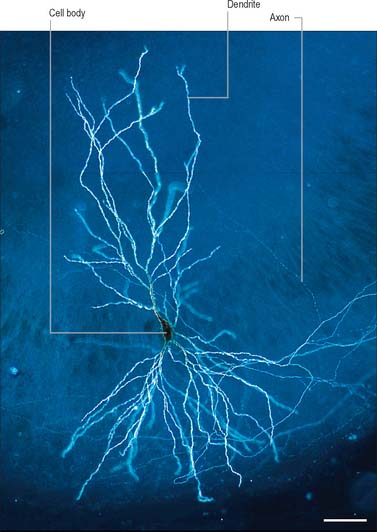
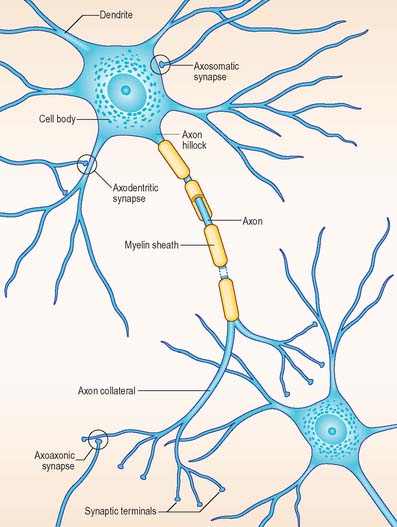
The huge complexity of the nervous system reflects the fact that individual neurones may make synaptic contact with hundreds, or even thousands, of other neurones via profuse axonal collateralization and dendritic branching (arborization). The latter is exemplified by the extensive dendritic field of the cerebellar Purkinje cell, which is traversed by thousands of axons, making synaptic contact as they pass. At the level of the individual neurone, competing incoming excitatory and inhibitory synaptic potentials are summated in time (temporal summation) and between synapses (spatial summation). If the postsynaptic neurone is depolarized above a certain threshold it fires action potentials which are conducted along the axon to the next target cells.
The nervous system is conventionally divided into two major parts, the central nervous system (CNS) and the peripheral nervous system (PNS). The CNS consists of the brain and spinal cord. The PNS consists of the cranial and spinal nerves and their ramifications, and certain groupings of cell bodies that constitute the peripheral ganglia. Another convention divides the nervous system into somatic and autonomic components, with elements in both the CNS and PNS. The somatic nervous system controls the contraction of skeletal muscles. The autonomic nervous system, which may be subdivided into sympathetic and parasympathetic components, consists of neurones that innervate secretory glands, cardiac and smooth muscle and are therefore concerned primarily with control of the internal environment. Neurones in the wall of the gastrointestinal tract form the enteric nervous system, and are capable of sustaining local reflex activity that is independent of the CNS (Ch. 73).
CENTRAL NERVOUS SYSTEM
The brain and spinal cord (see Fig. 43.1A) contain the great majority of neuronal cell bodies in the nervous system. In many parts of the CNS, neuronal cell bodies are grouped together and are, more or less, segregated from axons. The generic term for such collections of cell bodies is grey matter. Small aggregations of neuronal cell bodies, which usually share a common functional role, are termed nuclei. Neuronal dendrites and synaptic interactions are mostly confined to grey matter. Axons tend to be grouped together to form white matter, so called because axons are often ensheathed in myelin which confers a paler colouration. Axons that pass between similar origins or destinations within the CNS tend to run together in defined pathways, or tracts. These often cross the midline (decussate), which means that one half of the body is, in many respects, controlled by, and sends information to, the opposite side of the brain.
Some groups of neurones in the spinal cord and brain stem that subserve similar functions are organized into longitudinal columns. The neurones in these columns may be concentrated into discrete, discontinuous nuclei in some areas, e.g. the cranial nerve nuclei of the brain stem, or they may form more or less continuous longitudinal bands, as in much of the spinal cord (Fig. 15.3). Efferent neurones constitute three such columns. The axons of motor neurones in the somatic motor column innervate muscles derived from head somites. The two other columns are related to specialized features of head morphology: neurones in the branchial motor column innervate muscles derived from the wall of the embryonic pharynx (branchial muscles) and the visceral motor column supplies preganglionic parasympathetic fibres to glands and visceral smooth muscle. Four longitudinal cell columns are related to sensory functions. The general somatic sensory column essentially deals with general sensory information from the head; special somatic sensory neurones are related to the special senses and receive vestibular and auditory input; general visceral sensory neurones deal with information from widespread and varied visceral sensory endings; special visceral sensory neurones are related to the special sense of taste.
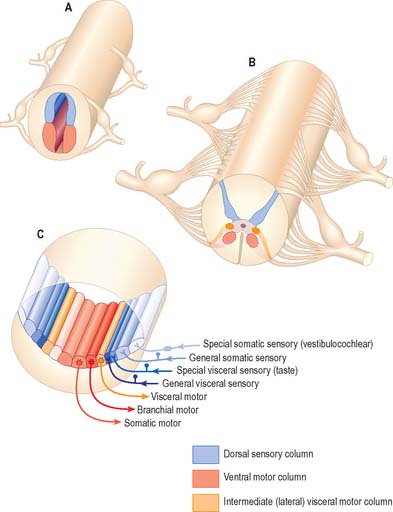
Fig. 15.3 Arrangement of sensory and motor cell columns in the spinal cord and brain stem. In each of parts A, B and C, dorsal is towards the top of the diagram. A shows the organization of the primitive spinal cord with a dorsal sensory column, a ventral motor column, and segmentally arranged dorsal and ventral nerve roots. B depicts the adult thoracic spinal cord with sensory and somatic motor columns and an additional intermediate (lateral) visceral motor column. C indicates the arrangement of multiple longitudinal columns in the brain stem, where the motor column is now subdivided into three, and the sensory column into four. For further information about the embryological aspects of the early nervous system consult Ch. 24. Consult also Fig 19.1.
To provide the energy required by constant neuronal activity, the CNS has a high metabolic rate and a rich blood supply (Ch. 17). A blood–brain barrier controls the neuronal environment and imposes severe restrictions on the types of substances that can pass from the bloodstream into nervous tissue (Ch. 3).
SPINAL CORD
The spinal cord lies within the vertebral column, in the upper two-thirds of the vertebral canal, and is continuous rostrally with the medulla oblongata of the brain stem (Ch. 43). For the most part, the spinal cord receives afferent input from, and controls the functions of, the trunk and limbs. Afferent and efferent connections between the periphery and the spinal cord travel in 31 pairs of segmentally arranged spinal nerves that attach to the cord as a linear series of dorsal and ventral rootlets. Adjacent groups of rootlets unite to form dorsal and ventral roots that combine to form the spinal nerves proper (Fig. 15.4). The dorsal and ventral roots are functionally distinct. Dorsal roots carry primary afferent nerve fibres from neuronal cell bodies located in dorsal root ganglia, whereas ventral roots carry efferent fibres from neuronal cell bodies located in the spinal grey matter.
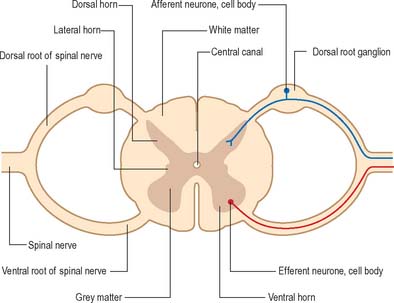
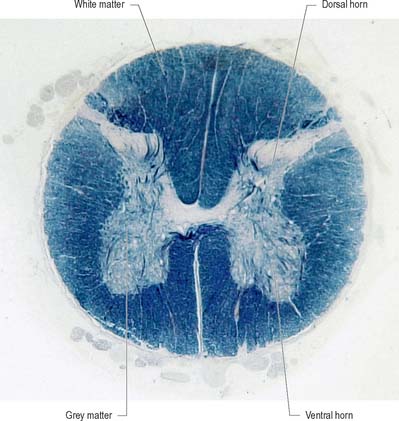
Internally, the spinal cord consists of a central core of grey matter surrounded by white matter. The grey matter is configured in a characteristic H, or butterfly, shape that has projections known as dorsal and ventral horns (Fig. 15.5). In general, neurones situated in the dorsal horn are primarily concerned with sensory functions whilst those in the ventral horn are mostly associated with motor activities. At certain levels of the spinal cord a small lateral horn is additionally present, marking the location of the cell bodies of preganglionic sympathetic neurones. The central canal, a vestigial component of the ventricular system, lies at the centre of the spinal grey matter and runs the length of the cord. The white matter of the spinal cord consists of ascending and descending tracts that link spinal cord segments to one another and the spinal cord to the brain.
BRAIN
The brain (encephalon) lies within the cranium (Ch. 26). The brain receives information from, and controls the activities of, the trunk and limbs mainly through connections with the spinal cord. It also possesses 12 pairs of cranial nerves through which it communicates mostly with structures of the head and neck. The brain is divided into major regions on the basis of ontogenetic growth and phylogenetic principles (Fig. 15.6, Fig. 15.7, Fig. 15.8). Ascending in sequence from the spinal cord, the principal divisions are the rhombencephalon or hindbrain, the mesencephalon or midbrain, and the prosencephalon or forebrain.
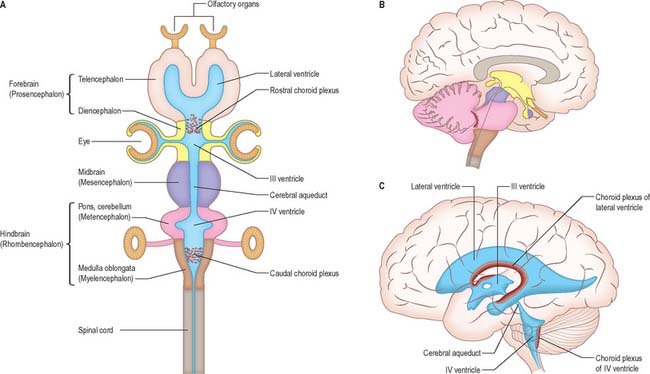
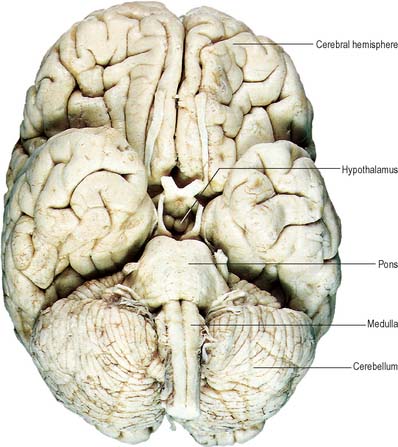
Fig. 15.7 Base of the brain, showing major divisions and cranial nerves.
(Figure enhanced by B Crossman, from a photograph by Kevin Fitzpatrick on behalf of GKT School of Medicine, London.)
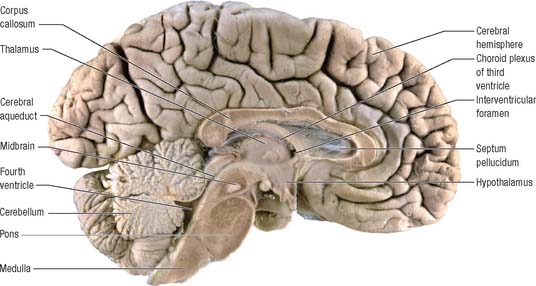
The prosencephalon is subdivided into the diencephalon and the telencephalon. The diencephalon equates mostly to the thalamus and hypothalamus, but also includes the smaller epithalamus and subthalamus. The telencephalon is mainly composed of the two cerebral hemispheres or cerebrum. The diencephalon is almost completely embedded in the cerebrum and is therefore largely hidden from the exterior. The human cerebrum constitutes the major part of the brain. It occupies the anterior and middle cranial fossae and is directly related to the cranial vault. It consists of two cerebral hemispheres. The surface of each hemisphere is convoluted into a complex pattern of ridges (gyri) and furrows (sulci). Internally, each hemisphere has an outer layer of grey matter, the cerebral cortex, beneath which lies a thick mass of white matter (Fig. 15.9). One of the most important components of the cerebral white matter, the internal capsule (see Fig. 15.11), contains nerve fibres which pass to and from the cerebral cortex and lower levels of the neuraxis. Several large nuclei of grey matter, usually referred to as the basal ganglia, are partly embedded in the subcortical white matter. Connections between corresponding areas of the two sides of the brain cross the midline within commissures. By far the largest commissure is the corpus callosum, which links corresponding regions of the two cerebral hemispheres.
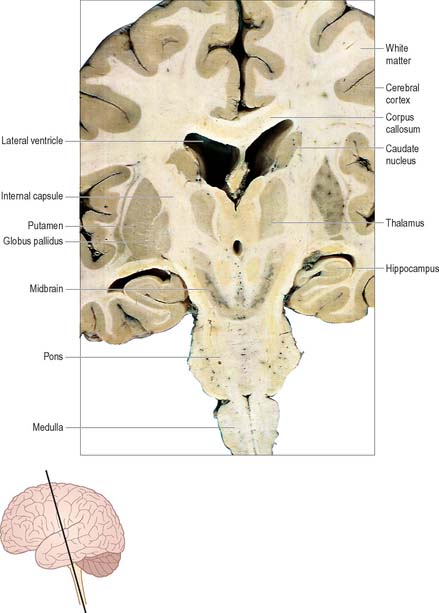
During prenatal development, the walls of the neural tube thicken greatly, but never completely obliterate the central lumen. Although the latter remains in the spinal cord as the narrow central canal, it becomes greatly expanded in the brain to form a series of interconnected cavities called the ventricular system (Ch. 16). In two regions, the fore- and hindbrains, parts of the roof of the neural tube do not generate nerve cells but become thin, folded sheets of highly vascular secretory tissue, the choroid plexuses. These secrete cerebrospinal fluid which fills the ventricles. The cavity of the rhombencephalon becomes expanded to form the fourth ventricle, which lies dorsal to the pons and upper half of the medulla. Caudally, the fourth ventricle is continuous with a canal in the caudal medulla and, through this, with the central canal of the spinal cord. The fourth ventricle is continuous with the subarachnoid space through three openings in its roof (the foramina of Luschka and of Magendie). At its rostral extent, the fourth ventricle is continuous with a narrow channel, the cerebral aqueduct, which passes through the midbrain. The rostral end of the cerebral aqueduct opens out into the third ventricle, a narrow, slit-like, midline cavity which is bounded laterally by the diencephalon. At the rostral end of the third ventricle, a small aperture on each side leads into the large lateral ventricle which is located within each cerebral hemisphere (see Fig. 15.6C).
OVERVIEW OF ASCENDING SENSORY PATHWAYS
Sensory modalities are conventionally described as being either special senses or general senses. The special senses are olfaction, vision, taste, hearing and vestibular function. Afferent information is encoded by highly specialized sense organs and transmitted to the brain in cranial nerves I, II, VII, VIII and IX (Ch. 25).
The general senses include touch, pressure, vibration, pain, thermal sensation and proprioception (perception of posture and movement). Stimuli from the external and internal environments activate a diverse range of receptors in the skin, viscera, muscles, tendons and joints (Ch. 3
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


