The cell cycle is governed by a series of checkpoints that determine the timing of each step in mitosis. In addition, checkpoints monitor and control the accuracy of DNA synthesis as well as the assembly and attachment of an elaborate network of microtubules that facilitate chromosome movement. If damage to the genome is detected, these mitotic checkpoints halt cell cycle progression until repairs are made or, if the damage is excessive, until the cell is instructed to die by programmed cell death (a process called apoptosis).
During G1, each cell contains one diploid copy of the genome. As the process of cell division begins, the cell enters S phase, the stage of programmed DNA synthesis, ultimately leading to the precise replication of each chromosome’s DNA. During this stage, each chromosome, which in G1 has been a single DNA molecule, is duplicated and consists of two sister chromatids (see Fig. 2-8), each of which contains an identical copy of the original linear DNA double helix. The two sister chromatids are held together physically at the centromere, a region of DNA that associates with a number of specific proteins to form the kinetochore. This complex structure serves to attach each chromosome to the microtubules of the mitotic spindle and to govern chromosome movement during mitosis. DNA synthesis during S phase is not synchronous throughout all chromosomes or even within a single chromosome; rather, along each chromosome, it begins at hundreds to thousands of sites, called origins of DNA replication. Individual chromosome segments have their own characteristic time of replication during the 6- to 8-hour S phase. The ends of each chromosome (or chromatid) are marked by telomeres, which consist of specialized repetitive DNA sequences that ensure the integrity of the chromosome during cell division. Correct maintenance of the ends of chromosomes requires a special enzyme called telomerase, which ensures that the very ends of each chromosome are replicated.
The essential nature of these structural elements of chromosomes and their role in ensuring genome integrity is illustrated by a range of clinical conditions that result from defects in elements of the telomere or kinetochore or cell cycle machinery or from inaccurate replication of even small portions of the genome (see Box). Some of these conditions will be presented in greater detail in subsequent chapters.
Clinical Consequences of Abnormalities and Variation in Chromosome Structure and Mechanics
Medically relevant conditions arising from abnormal structure or function of chromosomal elements during cell division include the following:
• A broad spectrum of congenital abnormalities in children with inherited defects in genes encoding key components of the mitotic spindle checkpoint at the kinetochore
• A range of birth defects and developmental disorders due to anomalous segregation of chromosomes with multiple or missing centromeres (see Chapter 6)
• A variety of cancers associated with overreplication (amplification) or altered timing of replication of specific regions of the genome in S phase (see Chapter 15)
• Roberts syndrome of growth retardation, limb shortening, and microcephaly in children with abnormalities of a gene required for proper sister chromatid alignment and cohesion in S phase
• Premature ovarian failure as a major cause of female infertility due to mutation in a meiosis-specific gene required for correct sister chromatid cohesion
• The so-called telomere syndromes, a number of degenerative disorders presenting from childhood to adulthood in patients with abnormal telomere shortening due to defects in components of telomerase
• And, at the other end of the spectrum, common gene variants that correlate with the number of copies of the repeats at telomeres and with life expectancy and longevity
By the end of S phase, the DNA content of the cell has doubled, and each cell now contains two copies of the diploid genome. After S phase, the cell enters a brief stage called G2. Throughout the whole cell cycle, the cell gradually enlarges, eventually doubling its total mass before the next mitosis. G2 is ended by mitosis, which begins when individual chromosomes begin to condense and become visible under the microscope as thin, extended threads, a process that is considered in greater detail in the following section.
The G1, S, and G2 phases together constitute interphase. In typical dividing human cells, the three phases take a total of 16 to 24 hours, whereas mitosis lasts only 1 to 2 hours (see Fig. 2-8). There is great variation, however, in the length of the cell cycle, which ranges from a few hours in rapidly dividing cells, such as those of the dermis of the skin or the intestinal mucosa, to months in other cell types.
Mitosis
During the mitotic phase of the cell cycle, an elaborate apparatus ensures that each of the two daughter cells receives a complete set of genetic information. This result is achieved by a mechanism that distributes one chromatid of each chromosome to each daughter cell (Fig. 2-9). The process of distributing a copy of each chromosome to each daughter cell is called chromosome segregation. The importance of this process for normal cell growth is illustrated by the observation that many tumors are invariably characterized by a state of genetic imbalance resulting from mitotic errors in the distribution of chromosomes to daughter cells.
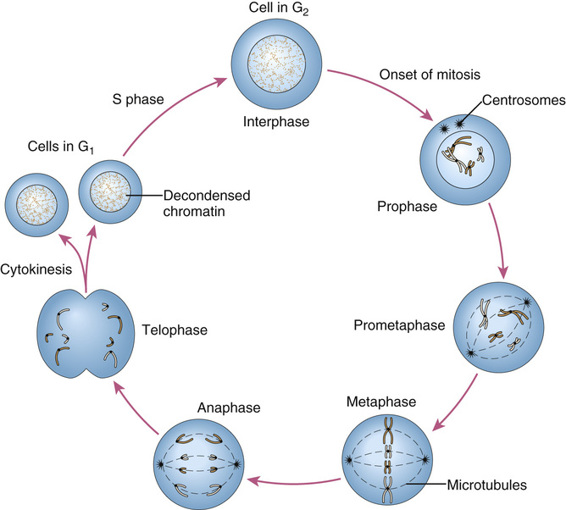
The process of mitosis is continuous, but five stages, illustrated in Figure 2-9, are distinguished: prophase, prometaphase, metaphase, anaphase, and telophase.
• Prophase. This stage is marked by gradual condensation of the chromosomes, formation of the mitotic spindle, and formation of a pair of centrosomes, from which microtubules radiate and eventually take up positions at the poles of the cell.
• Prometaphase. Here, the nuclear membrane dissolves, allowing the chromosomes to disperse within the cell and to attach, by their kinetochores, to microtubules of the mitotic spindle.
• Metaphase. At this stage, the chromosomes are maximally condensed and line up at the equatorial plane of the cell.
• Anaphase. The chromosomes separate at the centromere, and the sister chromatids of each chromosome now become independent daughter chromosomes, which move to opposite poles of the cell.
• Telophase. Now, the chromosomes begin to decondense from their highly contracted state, and a nuclear membrane begins to re-form around each of the two daughter nuclei, which resume their interphase appearance. To complete the process of cell division, the cytoplasm cleaves by a process known as cytokinesis.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


