Chapter 7 Ocular Embryology
With the explosion of information from improved technology and human genome study, new information is reported daily about the processes that control cellular development, structure, and function. A number of growth factors have been identified that bind to receptor sites on target cells to control normal development by modulating proliferation, migration, and differentiation.1 These processes are at the basis of structure development. Cells from germ layers migrate to a specific location and then proliferate, forming the population of cells that will differentiate into the specific cell type necessary.
Development of Ocular Structures
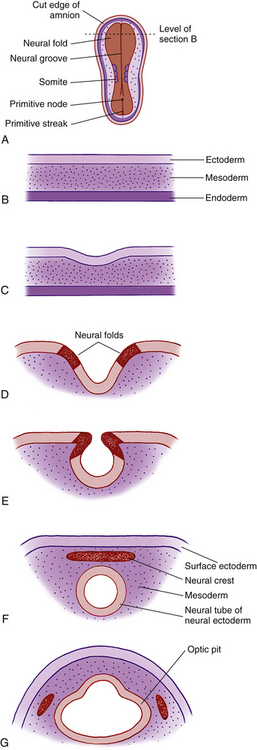
By the third week of embryonic development, the three primary germ layers—ectoderm, mesoderm, and endoderm—have formed the embryonic plate.2 (Of these three, only ectoderm and mesoderm will take part in the developing ocular structures.) A thickening in the ectoderm, visible on the dorsal surface of the embryo, forms the neural plate, which will give rise to the central nervous system, including ocular structures. A groove forms down the center of this plate at approximately day 18 of gestation, and the ridges bordering the groove grow into neural folds. As the groove expands, these folds grow toward one another and fuse to form the neural tube along the dorsal aspect of the embryo. Just before fusing, an area of cells on the crest of each of the neural folds separates from the ectoderm; these are neural crest cells. They form islands of cells within the mesoderm, which now surrounds the neural tube. The neural tube is formed on or near day 22.2 The tissue of the neural tube is now called neural ectoderm and the surface layer is now called surface ectoderm. Neural and surface ectoderm differ in anatomic location and in differentiation potentials (Box 7-1). Figure 7-1 illustrates these events.
BOX 7-1 Embryologic Derivation of Ocular Structures
Surface ectoderm gives rise to:
Neural ectoderm gives rise to:
Optic Pits
Indentations form in the inner surface of the neural tube on both sides of the forebrain region even before the tube is completely closed. These indentations are the optic pits. On approximately day 25, after the neural tube has closed, the optic pits expand forming lateral sac-shaped extensions, the optic vesicles.3 The cavity within the optic vesicle is continuous with the lumen of the neural tube. The surface of each vesicle expands until it comes in contact with surface ectoderm then gradually becomes separated from it by cells of neural crest origin and mesoderm.4
Neural crest cells and mesoderm collectively make up the mesenchyme, from which the connective tissue of the globe and orbit develop. Although most orbital connective tissue is derived from neural crest, determining whether a structure is of neural crest or mesodermal origin sometimes is difficult because mesodermal cells and neural crest cells appear similar cytologically.5 If the origin is uncertain, mesenchyme is cited as the germ layer.
As the optic vesicle evaginates, the tissue joining the vesicle to the neural tube constricts, forming the optic stalk (Figure 7-2). The cells lining the inner surface of this entire formation are ciliated, and the outer surface is covered by a thin basal lamina.3 The cavity of the optic stalk, as well as that of the optic vesicle, is continuous with the space that will become the third ventricle.
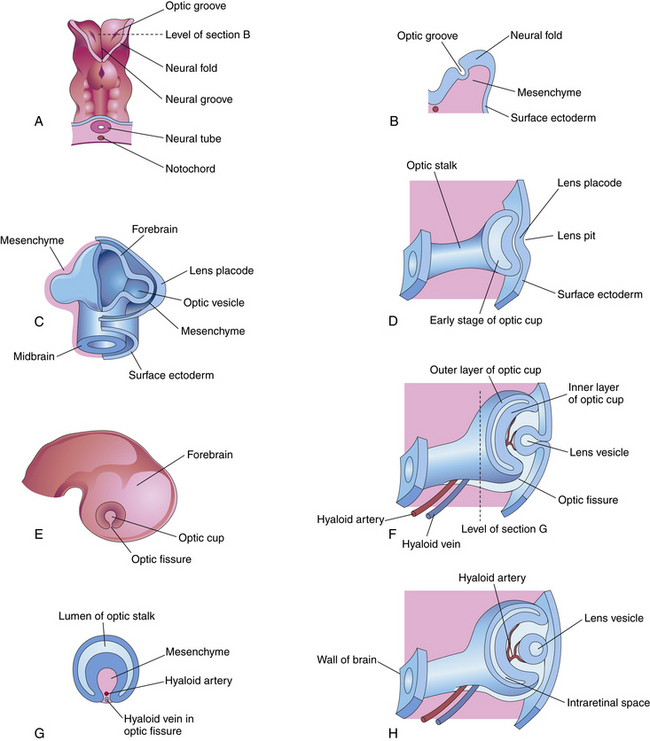
FIGURE 7-2 Early eye development.
(From Moore KL: Before we are born: essentials of embryology and birth defects, ed 5, Philadelphia, 1998, Saunders.)
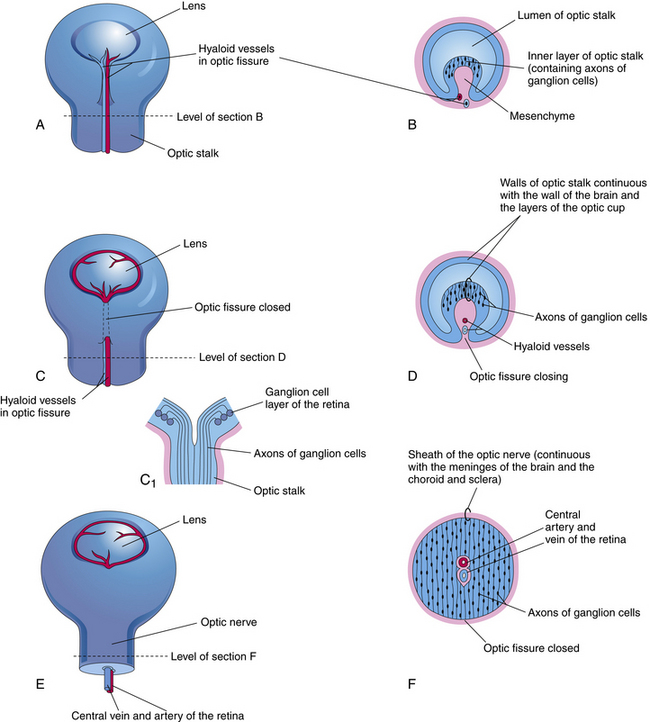
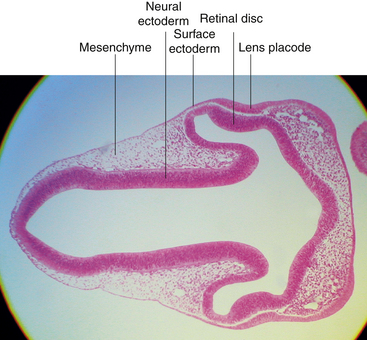
While the wall of the optic vesicle is in contact with surface ectoderm, it thickens and flattens to form the retinal disc.5 The lower wall of the optic vesicle and optic stalk begins to buckle and move inward toward the upper and posterior walls. This invagination forms a cleft, variously called the optic fissure, embryonic fissure, or fetal fissure. (It has also been called the “choroidal fissure,” but this name will be avoided because it may imply that the choroid is involved in the fissure, which it is not.) The inferior wall continues to move inward, pulling the anterior wall of the optic vesicle with it and placing the retinal disc in the approximate location of the future retina. The edges of the fissure grow toward one another and begin to fuse at 5 weeks; fusion starts at the center and proceeds anteriorly toward the rim of the optic cup and posteriorly along the optic stalk. Closure is complete at 7 weeks, forming the two layers of the optic cup and optic stalk6 (Figure 7-3). Mesenchyme enters the fissure and moves into the cavity of the developing optic cup.
Optic Cup, Lens, and hyaloid vessels
Optic Cup
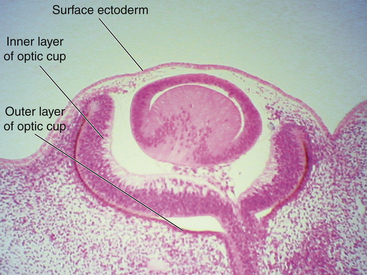
The optic cup at this stage of development is composed of two layers of cells (both neuroectodermal in origin) that are continuous with each other at the rim of the cup. The cells of the inner and outer layers of the optic cup are positioned apex to apex and are separated by the intraretinal space (see Figure 7-2, H), which, as the two layers approach each other, finally will become only a potential space. The outer layer of the optic cup will become the retinal pigment epithelium (RPE), the outer pigmented epithelium of the ciliary body, and the anterior iris epithelium. The inner layer will become the neural retina, the inner nonpigmented ciliary body epithelium, and the posterior iris epithelium.
Mesenchyme proliferates and migrates around the optic cup, and once the cells reach their destination, they proliferate and differentiate, contributing to the connective tissue of the eye and orbit. Neural crest cells will form corneal stroma and endothelium, uveal stroma and melanocytes, ciliary muscle, much of the sclera, the connective tissue and meningeal sheaths of the optic nerve, and the connective tissue of the lids, conjunctiva, and orbit. Vascular endothelium and striated muscle cells are formed by mesoderm.1
Clinical Comment: Coloboma
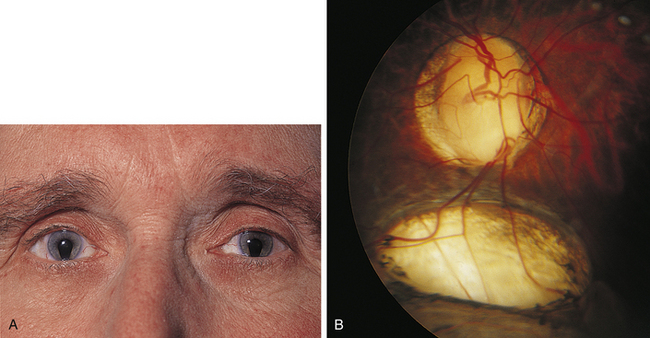
Incomplete closure of the optic fissure may affect the developing optic cup or stalk and the adult derivations of these structures, resulting in an inferior nasal defect in the optic disc, retina, ciliary body, or iris. This defect is called a coloboma and can vary from a slight notch to a large wedgelike defect. A large iris coloboma produces a keyhole-shaped pupil, although the remainder of the iris develops normally (Figure 7-4, A). When the coloboma is unilateral, the affected iris may have denser pigmentation than the opposite normal iris.7 Colobomas affecting the sensory retina and RPE also involve the choroid because its differentiation depends on an intact RPE layer.8 Bare sclera is seen in the area affected, with retinal vessels passing over the defect (Figure 7-4, B).
Induction
During embryologic development, formation and growth of structures depend on tissue differentiation and interactions among these tissues. Some structures will not develop unless they are near another developing area at a specific time. In some cases the two structures must actually come in contact; in others the structures must just be in proximity to each other, allowing biochemical signals to pass between them.9 The influence that one developing structure has on another is termed induction. It is likely that the mechanism of induction is not a single event but a series of separate steps that presumably occur on a biochemical level.10
Lens
Induction occurs between the developing optic cup and the developing lens, apparently through a reciprocal relationship.11 As the surface ectoderm comes in contact with the optic vesicle, the invagination of the optic cup begins (approximately day 27), and the surface ectoderm adjacent to the vesicle begins to thicken, forming the lens plate (lens placode) (Figures 7-5 and 7-6, A). This thickening is caused by an elongation of the ectodermal cells and by a regional increase in cell division.5 If the area of contact between the optic vesicle and surface ectoderm is less than normal, a perfectly formed but microphthalmic eye can result.12 Some investigators believe that transformation of the lens plate into the lens vesicle might be independent of direct contact with the optic vesicle.13 In addition to signals from the developing optic vesicle, complete lens differentiation might also depend on factors that inhibit lens formation in the ectoderm adjacent to the lens plate.14
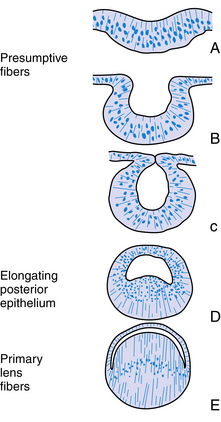
Cell division ceases in the center of the lens plate, forming a pit, and cell division accelerates in the periphery such that the lens plate invaginates rapidly.15 As invagination continues, the lens vesicle is formed and then separates from the surface ectoderm at approximately day 33.3,9 The lens vesicle is a hollow sphere composed of a single layer of cells, the apical surface of the cell lines the lumen, and the basal aspect is covered by a thin basal lamina; with the addition of more material, the basal lamina will become the lens capsule (Figure 7-6, B and C).
Once the lens vesicle is formed, the posterior epithelial cells adjacent to the future vitreous cavity elongate to fill in the lumen within the lens vesicle (Figure 7-6, D, E). In chick embryos if the lens is turned 180 degrees the epithelium that was once at the anterior of the lens vesicle is now adjacent to the developing optic cup and this is the epithelium that elongates filling in the lumen.16 The developing retina secretes biochemical mediators and is apparently the inducing factor for this elongation.15 The orientation of the lens is also influenced by the developing vitreous.12,17 The posterior epithelial cells become the primary lens fibers and form the embryonic nucleus at the center of the lens. This nucleus has no sutures. The fact that the posterior epithelium was used to form the embryonic nucleus accounts for the lack of an epithelial layer beneath the posterior lens capsule in the fully formed lens. The anterior epithelial cells remain in place, and the cells near the equator begin to undergo mitosis. Each new cell elongates anteriorly and posteriorly, forming secondary lens fibers that are laid down around the embryonic nucleus. The first layer of secondary fibers is completed by week 7.9 Secondary lens fibers continue to form and each layer surrounds the previous layer. The ends of the fibers meet in an upright Y-suture immediately posterior to the anterior epithelium and in an inverted Y-suture immediately anterior to the posterior capsule. These sutures are visible during the third month.18 The fetal nucleus contains the Y-sutures and all fibers formed before birth. If a line were drawn to connect the cellular nuclei within a lens fiber layer, an arcuate shape would be revealed. This configuration is called the lens bow (Figure 7-7).
Mitosis, cell elongation, and lens fiber formation continue throughout development and throughout life. The lens is initially spherical in shape but becomes more ellipsoid with additional fibers.16 The lens capsule is evident at 5 weeks, evolving from the basement membrane of the invaginating surface ectoderm and from secretions of the lens epithelium.18
Clinical Comment: Congenital Cataract
The spectrum of lens opacities that can result from problems during lens development range from pinpoint densities having no effect on vision to significant opacities causing extensive loss of vision.19 If the tissue near the developing lens fails to induce the lens fibers to elongate and pack together in an orderly way, the lens fibers will be misaligned, forming a cataract of the primary fibers. Interference with secondary lens fibers can lead to sutural cataracts.20
Viral infection affecting the mother during the first trimester often causes congenital malformations, including a cataract. The developing lens is vulnerable to the rubella virus (German measles) between the fourth and seventh week of development, when the primary fibers are forming. After this period the virus cannot penetrate the lens capsule and thus will not affect the lens. The cataract usually is present at birth but may develop weeks to months later because the virus can persist within the lens for up to 3 years. The opacity may be dense and opaque or it may be diffuse; the cataract may affect the nucleus only or may involve most of the lens.21
Hyaloid Arterial System
A branch of the internal carotid artery enters the optic cup through the fetal fissure to become the hyaloid artery during week 5.1 This vessel produces a highly branching network that fills the vitreous cavity and forms the posterior vascular tunic of the lens (posterior tunica vasculosa lentis), a vascular network covering the posterior lens (Figure 7-8). By the end of the second month, the hyaloid vasculature is fully formed.1 Branches near the lens equator anastomose with the annular vessel at the margin of the optic cup. The annular vessel sends loops forward onto the anterior surface of the lens to form the anterior vascular tunic of the lens (anterior tunica vasculosa lentis) during the seventh week1,22,23 (Figure 7-9). These vascular networks carry nutrients to the developing lens until production of the aqueous humor occurs. These vessels drain into a network located in the region that will become the ciliary body.3 The vessels of the hyaloid system cannot be identified as arterial or venous on the basis of their histologic makeup.22
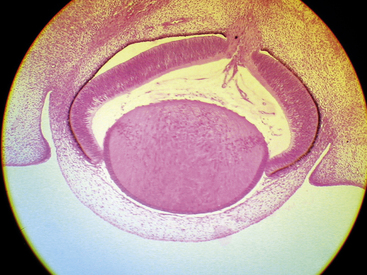
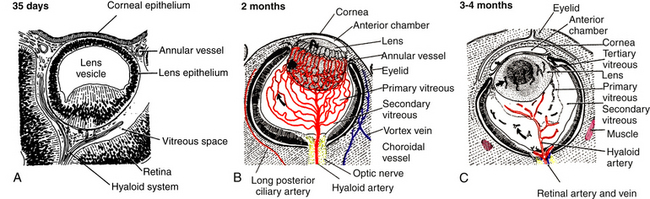
Glial cells on the surface of the optic cup form a conelike mass of tissue around the base of the hyaloid artery. These cells proliferate, forming a glial mantle around the arterial system. The hyaloid vasculature reaches its peak development during the third month1 and begins to atrophy during the fourth month, at the same time that the retinal vasculature is developing.1,24 By the seventh month, no blood flow is present in the hyaloid vasculature, which normally should be completely reabsorbed by birth.3 The extent of the degeneration of the glial tissue mass defines the extent of the adult physiologic optic cup.25
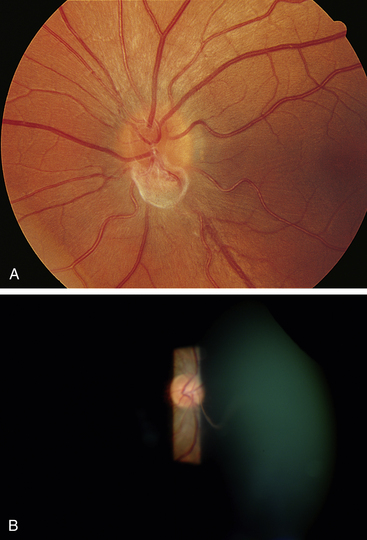
Clinical Comment: Bergmeister’s Papilla and Mittendorf’s Dot
Remnants of the hyaloid system often are seen clinically during examination of a patient’s ocular health. Glial tissue that persists on the nerve head is called Bergmeister’s papilla (Figure 7-10, A), and a pinpoint remnant of the hyaloid artery on the posterior surface of the lens is called Mittendorf’s dot. Rarely, a remnant of the entire hyaloid artery will be seen coursing through the vitreous from its attachment at the disc to the posterior lens (Figure 7-10, B).
Retinal Pigment Epithelium
Apposition of the two layers of the optic cup is essential for development of the retinal pigment epithelium,12 the first retinal layer to differentiate.25 Cellular structures and melanosomes begin to appear in the outer layer of the optic cup, and pigmentation of the retinal epithelium occurs at approximately week 3 or 4; this is the earliest pigmentation evident in the embryo3,4,26 (see Figure 7-8). After week 6, the RPE is one cell thick. The cells are cuboidal to columnar in shape, and the base of each cell is external toward the developing choroid and the apex internal toward the inner layer of the optic cup.27,28
Neural Retina
Between weeks 4 to 6, the cells of the inner layer of the optic cup (in the area that will become the neural retina) proliferate, and two zones are evident.1 The cells accumulate in the outer region, the proliferative zone or germinating zone. The inner marginal zone (of His) is anuclear. A thin lamina—the basement membrane of the inner layer of the optic cup and the precursor of the internal limiting membrane—separates the marginal zone from the vitreal cavity.18 At approximately week 7, cell migration occurs, forming the inner and outer neuroblastic layers, between which lies the transient fiber layer of Chievitz, a nucleus-free area.27,29 The formation of these two neuroblastic layers is complete during the third month. Differentiation of the neural retinal cells begins in the central retina and proceeds to the periphery.1
Ganglion cells and amacrine cells differentiate in the vitread portion of the inner neuroblastic layer.30 The ganglion cells migrate, forming a layer close to the basement membrane, and almost immediately send out their axonal processes, which become evident by week 8.18 Biomolecular agents guide axonal growth toward termination in the lateral geniculate nucleus.31,32 Müller cells, located rather centrally in the inner neuroblastic layer, develop at the same time. The bodies of the Müller and amacrine cells remain in the inner neuroblastic layer but move slightly sclerad.30
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


