Although the existence of nontuberculous mycobacteria (NTM) was recognized over a century ago, the microorganisms were originally thought to be contaminants or harmless colonizers. Increases in numbers of severely immunosuppressed patients, extensive utilization of invasive procedures, and more sensitive diagnostic tests have contributed to an increase in the isolation of the NTM from clinical samples (1, 2, 3, 4, 5and6,7). As a consequence, over the past four decades, the recognition and relative importance of NTM as a cause of human disease have increased dramatically. NTM are ubiquitous in nature, having been isolated from a variety of environmental sources, including dust, water, soil, domestic and wild animals, milk, and food (5,8, 9, 10, 11, 12, 13, 14, 15, 16, 17and18). More than 130 species are currently recognized. While many of these species are nonpathogenic, an increasing number, including Mycobacterium avium complex (MAC), M. kansasii, M. chelonae, M. fortuitum, M. mageritense, M. xenopi, M. lentiflavum, M. marinum, M. simiae, M. haemophilum, and M. genavense, have been associated with disease in normal and immunosuppressed hosts (1,2,4,19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29and30). A select number of these species have also been linked to healthcare-associated disease, including the M. fortuitum group, the M. chelonae/abscessus group (including M. immunogenum, M. chelonae, and M. abscessus [recently reclassified as M. abscessus subsp. abscessus and M. abscessus subsp. bolletii (31), and collectively referred to hereafter as M. abscessus group]), M. neoaurum, M. bacteremicum, M. kansasii, MAC, and M. xenopi.
MICROBIOLOGY
The slowly growing NTM, including those species usually associated with healthcare-associated diseases, grow well on the same types of media used for cultivation of M. tuberculosis complex (MTBC). Optimal incubation temperatures vary from 28°C (for species typically associated with cutaneous infections such as M. marinum, M. haemophilum, and M. chelonae) to 35°C to 37°C for most of the slowly growing NTM.
M. xenopi grows optimally at 42°C to 43°C but, with prolonged incubation, will grow at 37°C. Most species produce visible colonies on solid agar within 7 to 14 days. More than 60 species of rapidly growing mycobacteria (RGM) are recognized, with this number increasing rapidly with the primary use and availability of 16S ribosomal RNA (rRNA) gene sequencing for identification. The RGM are susceptible to the NaOH decontamination process performed on sputum to facilitate isolation of MTBC. They are nonfastidious microorganisms that produce mature, visible colonies on solid agar in 3 to 7 days. The microorganisms grow well at 30°C to 37°C on standard bacterial media, including 5% sheep’s blood and chocolate agar, and on media specifically formulated for mycobacterial species. Isolation of M. chelonae is optimal at an incubation temperature of 28°C to 32°C.
IDENTIFICATION
The reemergence of tuberculosis including multidrugresistant strains (MDR and XDR) as well as the heightened awareness of NTM as human pathogens has fueled an intense effort to develop rapid and accurate methods of identifying mycobacteria at the species level. Traditional identification schemes for slowly growing species utilize growth rates, pigment production, and biochemicals such as niacin production, urease, and catalase. Biochemical tests traditionally used in identifying the pathogenic RGM include the rapid (3-day) arylsulfatase reaction, iron uptake, nitrate reduction, and the ability to utilize mannitol, inositol, and/or citrate as carbon sources (32). Although these biochemical methods (utilized throughout the 1980s) have been replaced by molecular methods and high-performance liquid chromatography (HPLC), some laboratories report the NTM only to a group or complex level.
The decade of the 1990s saw identification of NTM by HPLC (33,34), while the last 10 years have seen a revolution of molecular-based tests, including commercial DNA probes, polymerase chain reaction (PCR) amplification followed by PCR restriction-enzyme analysis (PRA), 16S rRNA rpoB, hsp65, and other gene sequencing (35, 36, 37, 38, 39and40). HPLC methods for separation of mycolic acids have allowed the identification of some slowly growing mycobacterial species and some RGM groups or complexes, with a greater specificity and speed than traditional biochemical methods (33,34). Nonradioactive commercial probes are available (Accu-Probe, Gen-Probe) and are routinely utilized for identifying isolates of MTBC, MAC, M. kansasii, and M. gordonae. These probes offer excellent sensitivity and specificity, and because they can be used directly on broth cultures (usually the first medium to show growth), they have significantly reduced the time for final reporting of results (41,42). Currently, no commercial probes are available for identification of the RGM. Newest among the modern identification methods are adaptations of PCR technology for detection and identification of mycobacteria in clinical samples (43). Two different nucleic acid amplification techniques for assaying directly from sputum are now commercially available (Amplicor, Roche; and MTD test, Gen-Probe) and are approved for detection of M. tuberculosis. Currently, no systems are commercially available for direct detection of NTM from clinical specimens or for species identification of pure cultures, although several novel approaches have been published. One approach utilizes PCR to amplify the gene encoding the 16S rRNA. The amplified fragment is then analyzed by species-specific probes or partial nucleotide sequencing (39) for speciation. A second approach capitalizes on species-specific restriction fragment length polymorphisms (RFLPs) in a PCR-amplified segment of the 65 kDa heat shock protein gene (44, 45and46).
TYPING SYSTEMS
Typing systems for RGM have utilized a number of phenotypic and genotypic methods, including detailed species identification, heavy metal and antimicrobial susceptibility patterns, plasmid profiles, multilocus enzyme electrophoresis (MEE), pulsed-field gel electrophoresis (PFGE), and, more recently, random amplified polymorphic DNA (RAPD) PCR (47,48,49,50, 51and52). PFGE has proven to be a highly useful tool for strain typing of RGM. This method utilizes restriction endonucleases with rare recognition sites such as XbaI, DraI, and AsnI to generate a small series of large genomic restriction fragments (LRFs), the pattern of which is strain specific. Wallace et al. (48) described the use of PFGE to type M. chelonae and the M. abscessus group with three reference strains, 28 sporadic isolates, and 62 healthcare-associated isolates from 10 healthcare-associated outbreaks. LRF patterns satisfactory for comparison were achieved in 54% of the M. abscessus group and 90% of M. chelonae isolates by using the restriction endonucleases DraI, AsnI, XbaI, and SpeI. The sporadic isolates were all highly variable. Isolates from 5 of 10 outbreaks that gave satisfactory LRF patterns were identical. Strains that had been repetitively isolated from patients over periods of time ranging from 2 to 11 years demonstrated that LRF patterns were highly stable. Previous studies with M. fortuitum with this technique showed similar results, except that satisfactory LRF patterns were obtained with all strains studied (49,53). Environmental water isolates were identical (clonal) to some outbreak strains, indicating that water was the likely source of these past outbreaks. No human carrier or environmental nonwater sources have been identified as an outbreak source by this technique. PFGE is currently the most definitive epidemiologic tool available for comparing suspected outbreak strains of most isolates and species of RGM.
More recently, methods have been introduced for DNA stabilization, which prevent the DNA denaturation seen with PFGE with select strains of bacterial species. Application of one of these methods, the use of hydroxyurea in the running buffer, allows for quality PFGE patterns with the approximate 50% of the M. abscessus strains that produced broken DNA (50).
Another nucleic acid amplification technique, RAPD-PCR or arbitrary primer -PCR, has also been applied to the investigation of outbreaks of the M. abscessus group (50, 51and52). This technique offers the advantage of being simpler and unaffected by spontaneous lysis of the DNA sample during preparation for PFGE, as occurred previously with 50% of isolates of M. abscessus (48). Its major disadvantage is that fewer patterns are produced with each primer; hence, at least three primers that produce quality patterns are needed—a finding that likely reflects the closely related character of these strains (50,52).
PFGE has also been used to study clustering or pseudooutbreaks of slowly growing mycobacteria, including MTBC, M. xenopi (53,54), M. kansasii (55,56), M. simiae, (57, 58and59), and MAC (60, 61and62). Other fingerprint techniques used for slowly growing species include MEE with M. fortuitum (47), the M. abscessus group (63), M. simiae (64), and the use of hybridization with repetitive insertional elements for M. xenopi (53), M. kansasii (55), and M. avium (60). Previously, serotyping of MAC was used for strain typing (62) but currently has been replaced by molecular strain typing.
Recently, a commercial system from DiversiLab system (BioMerieux, Durham, NC), using repetitive elements interspersed throughout the genome, was introduced for strain typing of microorganisms including mycobacteria. The method was reported to be more rapid, required less sample size, and provided equivalent or better than standard RFLP for some species of mycobacteria (65).
Another method that has been evaluated recently with isolates of M. fortuitum and the M. abscessus/chelonae group is the enterobacterial repetitive intergenic consensus (ERIC) PCR. In an outbreak of the M. abscessus subsp. bolletii in Brazil recently, the ERIC PCR showed higher discrimination than PFGE for the M. abscessus group but less discriminatory power among isolates of M. fortuitum (66, 67and68).
Finally, multigene sequencing has recently been used to characterize isolates of Mycobacterium in hemodialysis water and may provide a reliable method of DNA strain typing (69).
EPIDEMIOLOGY
NTM are not reportable by law in most states, and thus, precise estimates of their incidence and prevalence are not available. Most NTM species, with the exception of MAC, are found in specific geographic areas. Overall, MAC is the most common NTM species recovered in the United States, followed by M. kansasii and the M. abscessus group (1). Although less frequently recovered in the United States, M. xenopi is the second most commonly isolated NTM species in England and Canada (70). In some areas of northern Europe, M. malmoense is second only to MAC (7). Although this species is rare in the United States, M. simiae is second to MAC in some cities in the southwestern United States (57,58,71,72). Tap water and biofilms in the pipes appear to be the major reservoirs for M. kansasii, M. xenopi, and M. simiae, and a reservoir for M. avium and M. intracellulare (73).
In contrast to surveys done in the late 1970s and early 1980s (1,70,74), more recent studies show that there are now more laboratory isolates of NTM in developed countries, especially MAC, than isolates of M. tuberculosis. The epidemiology of disease due to NTM has changed because of the improvement in laboratory recovery and identification of these species and the increased awareness of the clinician of these species as potential pathogens. The emergence of better antiretroviral therapies for human immunodeficiency virus (HIV) infection (acquired immunodeficiency syndrome [AIDS]) has resulted in a dramatic decline in the incidence of NTM disease in patients with far advanced disease. Among AIDS patients, MAC had been a common mycobacterial cause of opportunistic infection and a frequent cause of disseminated disease.
The RGM are the most commonly described and the most significant NTM for healthcare-associated epidemiology (75). Of the human diseases attributable to this group of microorganisms, over 90% are due to M. fortuitum, the M. abscessus group, and M. chelonae (2,13,21,75). These species readily survive nutritional deprivation and extremes of temperature. For example, most pathogenic species have been shown to grow and survive in distilled water, and they have been identified from soil, dust, domestic animals, and marine life (3,5,73,76,77). Multiple water sources have been identified, including tap water, municipal water, and aquariums (2,3,5,10). Mycobacteria have also been found in high numbers in biofilms on water-delivery devices, such as dental hand pieces (78). Similar biofilms may exist within bronchoscope channels, endoscope washers, ice machines, and water tanks, explaining the tendency of these devices to become colonized with mycobacteria. Biofilms are important not only because they enable bacteria to adhere and persist on artificial surfaces but also because they provide protection from the action of disinfectants (79, 80, 81and82).
PATHOGENESIS AND CLINICAL MANIFESTATIONS
The pathology of NTM infection can be identical to that of M. tuberculosis. Chronic inflammation, acute suppuration, nonnecrotic epithelioid tubercles, and caseation are all seen on histopathology. The coexistence of granulomatous and acute inflammation (so-called dimorphic inflammatory response) is not seen with tuberculosis but is commonly seen in cervical lymph nodes (83) and cutaneous disease (2,21,84) due to the NTM. Animal models to study the pathology of NTM have been difficult to develop, even when the animals are immunosuppressed (85,86).
Isolation of NTM in the laboratory may represent an environmental or laboratory contaminant, transient patient colonization, or true disease. In the absence of known environmental contamination, isolation of any NTM from a normally sterile site should be considered significant. Contamination of a skin wound with these microorganisms is rare, and even a single positive culture from this site generally indicates disease. Similarly, recovery of these microorganisms from cultures of lymph node specimens or blood is sufficient for establishing the diagnosis of nontuberculous lymphadenitis or disseminated disease, respectively.
In contrast, isolation of NTM from pulmonary specimens can be particularly difficult to evaluate. The American Thoracic Society last published criteria for the diagnosis of NTM pulmonary disease in 2007 (7). According to these criteria, a definitive diagnosis requires compatible clinical symptoms along with characteristic radiographic abnormalities, which are not attributable to any other cause. Multiple cultures of respiratory specimens are required to demonstrate persistent culture positivity. The microorganism must be grown from two acid-fast bacilli (AFB) specimens of sputum or from at least one specimen obtained from a normally sterile site such as a bronchial wash or bronchopulmonary tissue. The clinical syndromes most commonly associated with NTM infections and the microorganisms usually responsible are summarized in Table 39-1. The major risk factor for pulmonary NTM disease appears to be underlying bronchiectasis.
TABLE 39-1 Clinical Presentations of Nontuberculous Mycobacterial Species
Clinical Syndrome
Common Causes
Less Common Causes
Bronchopulmonary infection
M. avium complex
M. kansasii
M. abscessus group
M. xenopi
M. fortuitum group
M. chelonae
M. malmoense
M. immunogenum
M. szulgai
M. simiae
M. asiaticum
Lymphadenitis
M. avium complex
M. malmoense
M. abscessus group
M. fortuitum group
Disseminated disease
M. avium complex
M. chelonae
M. abscessus group
M. fortuitum group
M. haemophilum
M. genavense
M. kansasii
Skeletal and joint infection
M. marinum
M. avium complex
M. fortuitum group
M. abscessus group
M. kansasii
M. chelonae
M. haemophilum
M. goodii
Skin and soft tissue infection
M. marinum
M. fortuitum group
M. chelonae
M. abscessus group
M. ulcerans
M. haemophilum
M. smegmatis group
(M. goodii, M. wolinskyi)
M. mageritense
Note: M. abscessus group includes isolates now reclassified as M. abscessus subsp. abscessus and M. abscessus subsp. bolletii.
DESCRIPTION OF COMMUNITY-ACQUIRED INFECTIONS
Rapidly Growing Mycobacteria
Cutaneous Disease RGM most commonly cause posttraumatic and postsurgical skin and soft tissue infections but can also cause lymphadenitis, keratitis, suppurative arthritis, osteomyelitis, endocarditis, peritonitis, bacteremia, and disseminated disease (21,51,87, 88, 89and90,91,92, 93, 94, 95, 96, 97and98). In a review of 125 cases of infection due to RGM (2), 60% presented with cutaneous manifestations, half of which were due to penetrating trauma. The usual pathogens in this setting are M. fortuitum, the M. abscessus group, and the former M. fortuitum third biovariant complex (2,99). (The major species within the M. fortuitum third biovariant complex include M. porcinum, M. houstonense, M. senegalense, and less commonly, M. boenickei, M. brisbanense, and M. neworleansense.) Patients with the M. fortuitum group and the M. abscessus group are generally healthy, and druginduced immune suppression is not considered a risk factor in contrast to those patients with M. chelonae in which corticosteroid usage is a major risk factor.
Infections are typically chronic and may heal spontaneously or after surgical debridement. Even without medical intervention, the lesions usually remain well localized. Infections typically present as cellulitis with acute and chronic inflammation, which may form ulcers or sinus tracts with serous, watery drainage (2,51,84,100,101).
Disseminated Disease Disseminated disease due to RGM is typically related to immunosuppression, particularly corticosteroid therapy (91,92,93). Dissemination primarily occurs in patients with M. chelonae and, to a lesser degree, with M. abscessus. These patients typically have no history of trauma but present with multiple draining skin lesions. Infections with M. chelonae and the M. abscessus group have been described in solid organ transplant patients, including renal, heart, and lung transplant patients as well as patients with rheumatoid arthritis or other autoimmune disorders on long-term, low-dose corticosteroids (2,21,93,102). In one series of renal transplant patients, 10 patients with M. chelonae infections (including both M. chelonae and the M. abscessus group) were identified over a 6-year period from four hospitals (103).
Pulmonary Disease Chronic pulmonary disease may occur with RGM, especially in older women with bronchiectasis. The M. abscessus group accounts for >80% of these cases. Pulmonary infection is usually chronic, insidious, and slowly progressive. Lung infection may also be associated with MAC, and similarities between patients with MAC and those with the M. abscessus group suggest a common pathogenicity or host susceptibility defect (7). Patients may have minimal symptoms of cough or fatigue for many years, and subtle changes on high-resolution computed tomography scanning or subtle deterioration in pulmonary function may be the only markers of disease progression. Clinically, these patients are older and typically present with bilateral nodular interstitial disease associated with cylindrical bronchiectasis (2,21,75,104). Their presentation appears identical to that in women with MAC lung disease.
Patients with cystic fibrosis may also be involved with increasing frequency, and M. abscessus appears to be more frequent than MAC in this setting. Primary risk factors for susceptibility to NTM lung infection include bronchiectasis and chronic recurrent airway and parenchymal infections with other microorganisms (7).
Other RGM including M. chelonae, the M. smegmatis group, and M. fortuitum infrequently may be associated with pulmonary disease in such underlying disorders as achalasia or lipoid pneumonia (7).
Hypersensitivity pneumonitis has been observed among factory workers working with metalworking fluids contaminated with a newly described species, M. immunogenum (105), and among users of poorly maintained indoor hot tubs in association with MAC (106,107).
Bone and Joint Infection
Osteomyelitis may follow open bone fractures, puncture wounds, and hematogenous dissemination. The most frequent pathogens in this setting are members of the M. fortuitum group, although two newly described species in the M. smegmatis group—M. goodii and M. wolinskyi—may also be involved (7,108).
Slowly Growing Nontuberculous Mycobacteria
Among the slowly growing NTM, M. malmoense, M. kansasii, MAC, M. xenopi, and M. simiae can all occasionally cause community-acquired pulmonary or extrapulmonary disease in HIV-negative patients (4,7). MAC, M. kansasii, M. haemophilum, and M. genavense are the usual causes of infections including disseminated disease in HIV-infected persons. However, the slowly growing species are less frequent causes of healthcare-associated disease than the RGM (4,7). Detailed descriptions of the clinical disease associated with these microorganisms are reported elsewhere (4,7,13).
Previous studies in different sections of one hospital in Madrid, Spain, have shown identical strains of MAC in clinical samples—21/23 (91%) of urine isolates and 5/19 (26%) of respiratory isolates by PFGE and other typing methods such as hybridization with IS1245 (109). The investigation hypothesized that contamination of clinical samples with an environmental strain was the most likely cause since none of the patients with this strain had disease (109). A similar study in California examined potable water as a possible source of MAC infection in both AIDS and non-AIDS patients. The investigation revealed that the MAC isolates from potable water in three homes, two commercial buildings, one reservoir, and eight hospitals had varying degrees of genetic relatedness to 19 clinical isolates from 17 patients. Hospitals had the highest incidence (93%) of MAC isolates. Aronson et al. stated that the large number of isolates found in hospital water with a close genetic relationship to patient isolates suggested the possibility of a healthcare-associated spread of MAC to immunocompromised patients, especially AIDS patients, although a prospective epidemiologic study was not performed (110).
DESCRIPTION OF HEALTHCARE-ASSOCIATED INFECTIONS
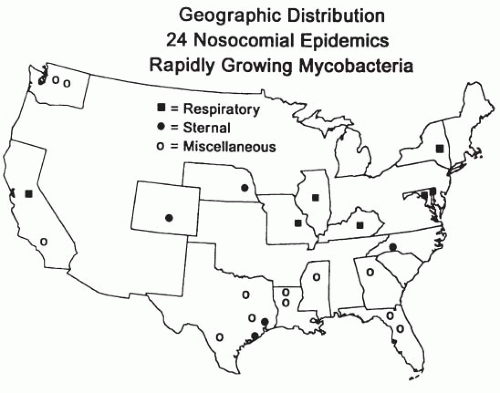
Healthcare-associated mycobacterial infections (almost exclusively due to RGM) have been recognized for more than 25 years and remain relatively common. They have been most often recognized primarily as causes of surgical site infections and postinjection abscesses; however, they have also been reported to cause catheter-related infections, dialysis-related infections, bronchoscope and endoscope contamination, and most recently, infections resulting from cosmetic procedures including plastic surgery, liposuction, and a healthcare-related practice involving subcutaneous injections of minute quantities of various drugs called mesotherapy (47,48,111,112,113,114,115). For outbreaks and sporadic reports of infections due to RGM, there is a strong geographic relationship to the Gulf Coast and southeastern states in the United States. Figure 39-1 demonstrates the focal geography of some of the early outbreaks reported in the United States. Recently, several outbreaks of NTM have occurred in South and Central America (66,116,117,118, 119, 120and121).
Surgical Site Infections
Outbreaks of mycobacterial surgical site infections were first recognized in 1975 to 1976 with the report of four such outbreaks (122, 123and124). These reports were followed in the 1980s by at least 14 additional outbreaks (125, 126, 127, 128, 129and130). Outbreaks have been described involving cardiothoracic surgery, plastic surgery, augmentation mammaplasty, and arthroplasty (122, 123, 124, 125, 126, 127, 128, 129and130). A summary of the major healthcare-associated outbreaks due to RGM is given in Table 39-2. Following recognition of epidemic surgical site infections, it became apparent that most surgical site infections due to RGM are sporadic (21,47,125,131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142and143). Such infections with RGM have been described following vascular surgery, oophorectomy, neurosurgery, corneal surgery, the insertion of middle ear tubes, biopsy procedures, and plastic surgery, including procedures such as face-lifts and liposuction (94,96,136,137,140,143, 144, 145, 146, 147, 148, 149and150). It is unclear whether there is a predisposition for certain types of surgery; however, more than 60% of surgical site infections due to RGM reported in the 1980s were reported after cardiac surgery (122,126,129,134) or augmentation mammaplasty (125,137,145). In the past 10 years, however, these latter surgeries have been replaced in incidence by cosmetic surgical procedures (111,116,117,118,120,151).
FIGURE 39-1 The geographic focality of healthcare-associated outbreaks due to rapidly growing mycobacteria.
Cardiothoracic Surgery
A review of RGM isolates associated with cardiac surgery was published in 1989 (47). This study evaluated isolates from eight cardiac surgery outbreaks, as well as 45 sporadic isolates. Disease isolates were recovered from sternal wounds, donor vein graft sites, blood, and artificial valves. The isolates included M. fortuitum, the M. abscessus group, and the M. smegmatis group (47). Several years later, the development of DNA fingerprinting for M. fortuitum and the M. abscessus group permitted better evaluation of these outbreaks (48,49,50, 51and52). The first reported cardiac surgery-associated outbreak occurred in 1976 in North Carolina. Nineteen cases of disease due to M. abscessus occurred over a 10-week period, but no source was identified. Five (26%) patients died of their disease. In a second similar outbreak in Colorado, 10 of 75 cardiac surgery patients developed infections with M. fortuitum. M. fortuitum was recovered from a settling plate in the operating room (122), but subsequent molecular studies using PFGE showed that the environmental strain differed from the disease strain (***53a).
The best clue to the potential reservoir for these outbreaks was provided in a later outbreak from Texas involving both M. fortuitum and the M. abscessus group. An isolate of M. fortuitum with a DNA fingerprint identical to that of the outbreak strain was isolated from the tap water in the operating room, ice water used to cool the cardioplegia solution, ice machines, and municipal water coming into the hospital. An identical strain was recovered from patients with several types of noncardiac surgical site infections. In this same outbreak, RAPD-PCR showed M. abscessus isolated from hospital ice water used to cool the cardioplegia solution and a pair of surgical scissors was identical to some disease isolates (47,52,***53a,126). This investigation was the first major study to identify water (in this case as ice used for surgical purposes) as the major reservoir for the microorganism. Another unreported outbreak in Texas also helped to clarify the role of water as a reservoir for RGM. In this outbreak, PFGE demonstrated the clonality of tap water and case isolates (Fig. 39-2). Several cases of perivalvular infection occurred following contamination of commercial porcine valves with M. chelonae (152,153).
Only gold members can continue reading. Log In or Register to continue