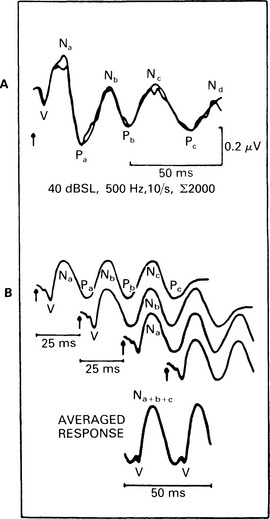
8 Middle-latency responses (MLR) recorded from the scalp have latencies in the range 10–50 ms. Some of the potentials are of myogenic origin, and most respond only to relatively high-intensity acoustic stimuli. However, the postauricular muscle response (PAM) has a relatively low threshold (Thornton 1975). Middle-latency neurogenic potentials are thought to originate in the thalamus and its primary projections, and possibly in the primary cortex. The middle-latency responses in man were described as long ago as 1958 by Geisler et al, and they have been studied by Goldstein & Rodman (1967) and Mendel & Goldstein (1969). Initially, they were called ‘early’ responses. However, they were renamed ‘middle-latency responses’ soon after the discovery of ABRs, which were classified as early-latency responses (Davis 1976). Both myogenic and neurogenic potentials occur in the time domain when the stimulus is of high intensity (Bickford et al 1964, Picton et al 1974). However, for low- and moderate-intensity sounds, there is substantial evidence of the neural origin of middle-latency response (Maast 1965). At least a partly neural origin of middle-latency response has been confirmed in the induction of muscle paralysis during anaesthesia. Wave morphology similar to MLR was obtained by Harker et al (1977), as was Pa potential by Kileny et al (1983). The sites of MLR generators have not yet been determined with certainty, but for the earlier MLR components No, Po, and Na, the medial geniculate ganglion and the thalamus have been suggested (Davis 1976). Generation of No, Po, and Na or SN10 has also been attributed to postsynaptic activity from the inferior colliculus in man (Hashimoto 1982). Evidence has been given for a primary cortical origin of a middle-latency auditory-evoked potential in laboratory animals, e.g. cats (Kaga et al 1980, Buchwald et al 1981). However, in man there is some uncertainty whether MLR components arise in the primary cortex. Parving et al (1980) also observed normal Pa in patients with auditory agnosia due to temporal lobe lesion. Kraus et al (1982) also observed normal Na and Pa in patients with unilateral temporal lobe lesion, but found elimination or reduction of the amplitudes in patients with bitemporal lesions. Also, generators of the Na component are more effectively activated by contralateral stimulation (Woods & Clayworth 1985). The post-auricular muscle response was first reported by Kiang et al (1963), who described it as a variable low-threshold response. Douek et al (1973) stressed the importance of recording from both sides simultaneously, as the response has a bilateral representation. Picton et al (1974) reported that the PAM response was highly variable, both within and among subjects. However, it was found that when using stimulation rate of 40/s (40 Hz), the main peaks become superimposed, interfere constructively, and produce a sinusoidal response (Galambos et al 1981). This ‘40-Hz’ MLR response can be used for threshold estimation (see Fig. 8.1).
Middle-latency responses (MLR)
INTRODUCTION
GENERATORS OF MIDDLE-LATENCY RESPONSES
METHODS
Technical aspects of stimulation and MLR recording
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Middle-latency responses (MLR)