The main factor determining the colour of skin is the degree of pigmentation produced by melanocytes in the basal layer of the epidermis. Melanocyte numbers are similar in all races. In darker skins the melanocytes produce more pigment. Melanins vary in colour from yellows to browns and blacks.
Sweat glands are distributed all over the skin except on the tympanic membranes, lip margins, nipples, inner surface of prepuce, glans penis and labia minora. The greatest concentration is in the thick skin of the palms and soles, and on the face. Sweat glands are coiled tubular structures that extend into the dermis and subcutaneous tissue. They are supplied by cholinergic fibres in sympathetic nerves. Apocrine glands are large, modified sweat glands confined to the axillae, areolae, periumbilical, genital and perianal regions; their ducts open into hair follicles or directly on to the skin surface. Their odourless secretion acquires a smell through bacterial action. They enlarge at puberty and undergo cyclic changes in relation to the menstrual cycle in females. They are supplied by adrenergic fibres in sympathetic nerves.
Sebaceous glands are small saccular structures in the dermis, where they open into the side of hair follicles. They also open directly on to the surface of the hairless skin of the lips, nipples, areolae, inner surface of prepuce, glans penis and labia minora. There are none on the palms or soles. They are particularly large on the face. Androgens act on these glands which have no motor innervation.
Hair and nails are a hard type of keratin; the keratin of the skin surface is soft keratin. Each hair is formed from the hair matrix, a region of epidermal cells at the base of the hair follicle, which extends deeply into the dermis and subcutaneous tissue. As the cells move up inside the tubular epidermal sheath of the follicle they lose their nuclei and become converted into the hard keratin hair shaft. Melanocytes in the hair matrix impart pigment to the hair cells. The change with age is due to decreasing melanocyte activity. An arrector pili muscle attached to the connective tissue of the base of the follicle passes obliquely to the upper part of the dermis. Contraction of this smooth muscle, with a sympathetic innervation, makes the hair ‘stand on end’, and squeezes the sebaceous gland that lies between the muscle and the hair follicle. Hair follicles are richly supplied by sensory nerves.
Nails consist of nail plates lying on nail beds on the dorsum of the terminal segment of fingers and toes. Compacted keratin-filled squames form the nail plate, which develops from epidermal cells deep to its proximal part. Here the nail plate is overlapped by the skin of the proximal nail fold. Blood vessels and sensory nerve endings are plentiful in the nail bed.
The arteries of the skin are derived from a tangential plexus in the subcutaneous connective tissue. Branches from this plexus form a subpapillary network in the dermis (Fig. 1.1). The veins have a similar arrangement to the arteries and arteriovenous anastomoses are abundant. From a meshwork of lymphatic capillaries in the papillary layer of the dermis, lymphatics pass inwards and then run centrally with the blood vessels. Cutaneous nerves carry afferent somatic fibres, mediating general sensation, and efferent autonomic (sympathetic) fibres, supplying smooth muscle of blood vessels, arrector pili muscles and sweat glands. Both free sensory nerve endings and several types of sensory receptors are present in the skin.
The proportionate surface area of the skin over different regions of the body can be estimated by the ‘rule of nines’ and this is useful in assessing the need for fluid replacement after burns. This rule is a guide to the size of body parts in relation to the whole: head 9%; upper limb 9%; lower limb 18%; front of thorax and abdomen 18%; back of thorax and abdomen 18%.
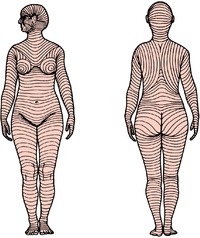
Tension lines of the skin, due to the patterns of arrangement of collagen fibres in the dermis, run as shown in Figure 1.2. They are often termed relaxed skin tension lines because they coincide with fine furrows present when the skin is relaxed. Wrinkle lines are caused by the contraction of underlying muscles; they do not always correspond to tension lines. Flexure lines over joints run parallel to tension lines. The cleavage lines originally described by Langer in 1861 on cadavers do not entirely coincide with the lines of greatest tension in the living. Incisions made along skin tension lines heal with a minimum of scarring (Fig. 1.3).
 |
| Figure 1.2 |
Superficial fascia
The skin is connected to the underlying bones or deep fascia by a layer of loose areolar connective tissue. This layer, usually referred to as superficial fascia, is of variable thickness and fat content. Flat sheets of muscles are also present in some regions. These include both skeletal muscles (platysma, palmaris brevis) and smooth muscles (subareolar muscle of the nipple, dartos, corrugator cutis ani). The superficial fascia is most distinct on the lower abdominal wall where it differentiates into two layers. Strong connective tissue bands traverse the superficial fasica binding the skin to the underlying aponeurosis of the scalp, palm and sole.
Deep fascia
The limbs and body wall are wrapped in a membrane of fibrous tissue, the deep fascia. It varies widely in thickness. In the iliotibial tract of the fascia lata, for example, it is very well developed, while over the rectus sheath and external oblique aponeurosis of the abdominal wall it is so thin as to be scarcely demonstrable and is usually considered to be absent. In other parts, such as the face and the ischioanal fossa, it is entirely absent. Where deep fascia passes directly over bone it is always anchored firmly to the periosteum and the underlying bone is described as being subcutaneous. In the neck, as well as the investing layer of deep fascia, there are other deeper fascial layers enclosing neurovascular structures, glands and muscles. Intermuscular septa are laminae of deep fascia which extend between muscle groups. Transverse thickenings of deep fascia over tendons, attached at their margins to bones, form retinaculae at the wrists and ankles and fibrous sheaths on the fingers and toes.
Ligaments
Ligaments are composed of dense connective tissue, mainly collagen fibres, the direction of the fibres being related to the stresses which they undergo. In general ligaments are unstretchable, unless subjected to prolonged strain. A few ligaments, such as the ligamenta flava between vertebral laminae and the ligamentum nuchae at the back of the neck, are made of elastic fibres, which enables them to stretch and regain their original length thereafter. Ligaments are usually attached to bone at their two ends.
Tendons
Tendons have a similar structure to collagenous ligaments, and attach muscle to bone. They may be cylindrical, or flattened into sheet-like aponeuroses. Tendons have a blood supply from vessels which descend from the muscle belly and anastomose with periosteal vessels at the bony attachment.
Synovial sheaths
Where tendons bear heavily on adjacent structures, and especially where they pass around loops or pulleys of fibrous tissue or bone and change the direction of their pull, they are lubricated by being provided with a synovial sheath. The parietal layer of the sheath is attached to the surrounding structures, the visceral layer is fixed to the tendon, and the two layers glide on each other, lubricated by a thin film of synovial fluid secreted by the lining cells of the sheath. The visceral and parietal layers join each other at the ends of their extent. Usually they do not enclose the tendon cylindrically; it is as though the tendon was pushed into the double layers of the closed sheath from one side. In this way blood vessels can enter the tendon to reinforce the longitudinal anastomosis. In other cases blood vessels perforate the sheath and raise up a synovial fold like a little mesentery—a vinculum—as in the flexor tendons of the digits (see Fig. 2.47C, p. 90).
Cartilage
Cartilage is a type of dense connective tissue in which cells are embedded in a firm matrix, containing fibres and ground substance composed of proteoglycan molecules, water and dissolved salts. There are three types of cartilage. The most common is hyaline cartilage which has a blue-white translucent appearance. Costal, nasal, most laryngeal, tracheobronchial, articular cartilage of typical synovial joints and epiphyseal growth plates of bones are hyaline cartilage.
Fibrocartilage is like white fibrous tissue but contains small islands of cartilage cells and ground substance between collagen bundles. It is found in intervertebral discs, the labrum of the shoulder and hip joints, the menisci of the knee joints and at the articular surface of bones which ossify in membrane (squamous temporal, mandible and clavicle). Both hyaline cartilage and fibrocartilage tend to calcify and they may even ossify in old age.
Elastic cartilage has a matrix that contains a large number of yellow elastic fibres. It occurs in the external ear, auditory (Eustachian) tube and epiglottis. Elastic cartilage never calcifies.
Fibrocartilage has a sparse blood supply, but hyaline and elastic cartilage have no capillaries, their cells being nourished by diffusion through the ground substance.
Muscle
There are three kinds of muscle—skeletal, cardiac and smooth—although the basic histological classification is into two types: striated and non-striated. This is because both skeletal and cardiac muscle are striated, a structural characteristic due to the way the filaments of actin and myosin are arranged. The term striated muscle, however, is usually taken to mean skeletal muscle. Smooth muscle, also known as visceral muscle, is non-striated. Smooth muscle also contains filaments of actin and myosin, but they are arranged differently. The terms ‘muscle cell’ and ‘muscle fibre’ are synonymous. Smooth muscle fibres have a single nucleus, cardiac muscle fibres have one or two nuclei and skeletal muscle fibres are multinucleated.
Smooth muscle consists of narrow spindle-shaped cells, usually lying parallel. They are capable of slow but sustained contraction. In tubes that undergo peristalsis they are arranged in longitudinal and circular fashion (as in the alimentary canal and ureter). In viscera that undergo a mass contraction without peristalsis (such as urinary bladder and uterus) the fibres are arranged in whorls and spirals rather than demonstrable layers. Contractile impulses are transmitted from one cell to another at sites called nexuses or gap junctions, where adjacent cell membranes lie unusually close together. Innervation is by autonomic nerves.
Cardiac muscle consists of broader, shorter cells that branch. Cardiac muscle is less powerful than skeletal muscle, but is more resistant to fatigue. Part of the boundary membranes of adjacent cells make very elaborate interdigitations with one another to increase the surface area for impulse conduction. The cells are arranged in whorls and spirals; each chamber of the heart empties by mass contraction. Although the heart has an intrinsic impulse generating and conduction system, the rate and force of contraction are influenced by autonomic nerves.
Skeletal muscle consists of long, cylindrical non-branching fibres. Individual fibres are surrounded by a fine network of connective tissue, the endomysium. Parallel groups of fibres are surrounded by less delicate connective tissue, the perimysium, to form muscle bundles or fasciculi. Thicker connective tissue, the epimysium, envelops the whole muscle. Neurovascular structures pass along the sheaths.
The orientation of individual skeletal muscle fibres is either parallel or oblique to the line of pull of the whole muscle. The range of contraction is long with the former arrangement, while the latter provides increased force of contraction. Sartorius is an example of a muscle with parallel fibres.
Muscles with an oblique disposition of fibres fall into several patterns:
• Unipennate muscles, where all the fibres slope into one side of the tendon, giving a pattern like a feather split longitudinally (e.g. flexor pollicis longus).
• Bipennate muscles, where muscle fibres slope into the two sides of a central tendon, like an intact feather (e.g. rectus femoris).
• Multipennate muscles, which take the form of a series of bipennate masses lying side by side (e.g. subscapularis), or of a cylindrical muscle within which a central tendon forms. Into the central tendon the sloping fibres of the muscle converge from all sides (e.g. tibialis anterior).
The attachment of a muscle, where there is less movement, is generally referred to as its origin, and the attachment, where there is greater movement, as its insertion. These terms are relative; which end of the muscle remains immobile and which end moves depends on circumstances and varies with most muscles. Simple usage of ‘attachment’ for both sites of fixation of a muscle avoids confusion and inaccuracy.
Movements are the result of the coordinated activity of many muscles, usually assisted or otherwise by gravity. Bringing the attachments of a muscle (origin and insertion) closer together is what is conventionally described as the ‘action’ of a muscle (isotonic contraction, shortening it). If this is the desired movement the muscle is said to be acting as a prime mover, as when biceps is required to flex the elbow. A muscle producing the opposite of the desired movement—triceps in this example—is acting as an antagonist; it is relaxing but in a suitably controlled manner to assist the prime mover. Two other classes of action are described: fixators and synergists. Fixators stabilize one attachment of a muscle so that the other end may move, e.g. muscles holding the scapula steady are acting as fixators when deltoid moves the humerus. Synergists prevent unwanted movement; the long flexors of the fingers pass across the wrist joint before reaching the fingers, and if finger flexion is the required movement, muscles that extend the wrist act as synergists to stabilize the wrist so that the finger flexors can act on the fingers. A muscle that acts as a prime mover for one activity can of course act as an antagonist, fixator or synergist at other times. Muscles can also contract isometrically, with increase of tension but the length remaining the same, as when the rectus abdominis contracts prior to an anticipated blow on the abdomen. Many muscles can be seen and felt during contraction, and this is the usual way of assessing their activity, but sometimes more specialized tests such as electrical stimulation and electromyography may be required.
Muscles have a rich blood supply. Arteries and veins usually pierce the surface in company with the motor nerves. From the muscle belly vessels pass on to supply the adjoining tendon. Lymphatics run back with the arteries to regional lymph nodes.
Embedded among the ordinary skeletal muscle cells are groups of up to about 10 small specialized muscle fibres that constitute the muscle spindles. The spindle fibres are held together as a group by a connective tissue capsule and are called intrafusal fibres (lying within a fusiform capsule), in contrast to ordinary skeletal muscle fibres which are extrafusal. Spindles act as a type of sensory receptor, transmitting to the central nervous system information on the state of contraction of the muscles in which they lie.
Skeletal muscle is supplied by somatic nerves through one or more motor branches which also contain afferent and autonomic fibres. The efferent fibres in spinal nerves are axons of the large α anterior horn cells of the spinal cord which pass to extrafusal fibres, and axons of the small γ cells which supply the spindle (intrafusal) fibres. The motor nuclei of cranial nerves provide the axons for those skeletal muscles supplied by cranial nerves.
The nerves supplying the ocular and facial muscles (third, fourth, sixth and seventh cranial nerves) contain no sensory fibres. Proprioceptive impulses are conveyed from the muscles by local branches of the trigeminal nerve. The spinal part of the accessory nerve and the hypoglossal nerve likewise contains no sensory fibres. Proprioceptive impulses are conveyed from sternoclei-domastoid and trapezius by branches of the cervical plexus, and from the tongue muscles probably by the lingual nerve (trigeminal).
Bone
Bone is a type of vascularized dense connective tissue with cells embedded in a matrix composed of organic materials, mainly collagen fibres, and inorganic salts rich in calcium and phosphate.
Macroscopically, bone exists in two forms: compact and cancellous. Compact bone is hard and dense, and resembles ivory. It occurs on the surface cortex of bones, being thicker in the shafts of long bones, and in the surface plates of flat bones. The collagen fibres in the mineralized matrix are arranged in layers, embedded in which are osteocytes. Most of these lamellae are arranged in concentric cylinders around vascular channels (Haversian canals), forming Haversian systems or osteons, which usually lie parallel to each other and to the long axis of the bone. Haversian canals communicate with the medullary cavity and each other by transversely running Volkmann’s canals containing anastomosing vessels. Cancellous bone consists of a spongework of trabeculae, arranged not haphazardly but in a very real pattern best adapted to resist the local strains and stresses. If for any reason there is an alteration in the strain to which cancellous bone is subjected there is a rearrangement of the trabeculae. The moulding of bone results from the resorption of existing bone by phagocytic osteoclasts and the deposition of new bone by osteoblasts. Cancellous bone is found in the interior of bones and at the articular ends of long bones. The organization of cancellous or trabecular bone is also basically lamellar but in the form of branching and anastomosing curved plates. Blood vessels do not usually lie within this bony tissue and osteocytes depend on diffusion from adjacent medullary vessels.
The medullary cavity in long bones and the interstices of cancellous bone are filled with red or yellow marrow. At birth all the marrow of all the bones is red, active haemopoiesis going on everywhere. As age advances the red marrow atrophies and is replaced by yellow, fatty marrow, with no power of haemopoiesis. This change begins in the distal parts of the limbs and gradually progresses proximally. By young adult life red marrow remains only in the ribs, sternum, vertebrae, skull bones, girdle bones and the proximal ends of the femur and humerus; these tend to be sites of deposition of malignant metastases.
The outer surfaces of bones are covered with a thick layer of vascular fibrous tissue, the periosteum, and the nutrition of the underlying bone substance depends on the integrity of its blood vessels. The periosteum is osteogenic, its deeper cells differentiating into osteoblasts when required. In the growing individual new bone is laid down under the periosteum, and even after growth has ceased the periosteum retains the power to produce new bone when it is needed, e.g. in the repair of fractures. The periosteum is united to the underlying bone by collagen (Sharpey’s) fibres, particularly strongly over the attachments of tendons and ligaments. Periosteum does not, of course, cover the articulating surfaces of the bones in synovial joints; it is reflected from the articular margins and blends with the capsule of the joint.
The single-layered endosteum that lines inner bone surfaces (marrow cavity and vascular canals) is also osteogenic and contributes to new bone formation.
One or two nutrient arteries enter the shaft of a long bone obliquely and are usually directed away from the growing end. Within the medullary cavity they divide into ascending and descending branches. Near the ends of bone they are joined by branches from neighbouring vessels and from periarticular arterial anastomoses. Cortical bone receives blood supply from the periosteum and from muscular vessels at their attachments. Veins are numerous and large in the cancellous red marrow bones (e.g. the basivertebral veins). Lymphatics are present, but scanty; they drain to the regional lymph nodes of the part.
Subcutaneous periosteum is supplied by the nerves of the overlying skin. In deeper parts the local nerves, usually the branches to nearby muscles, provide the supply. Periosteum in all parts of the body is very sensitive. Other nerves, probably vasomotor in function, accompany nutrient vessels into bone.
Bone develops by two main processes, intramembranous and endochondral ossification (ossification in membrane and cartilage). In general the bones of the vault of the skull, the face and the clavicle ossify in membrane, while the long bones of the skeleton ossify in cartilage.
In intramembranous ossification, osteoblasts simply lay down bone in fibrous tissue; there is no cartilage precursor. The bones of the skull vault, face and the clavicle develop in this way and the growth in the thickness of other bones (subperiosteal ossification) is also by intramembranous ossification.
In endochondral ossification a pre-existing hyaline cartilage model of the bone is gradually destroyed and replaced by bone. The majority of the bones of the skeleton, including the long bones, are formed in this way. The cartilage is not converted into bone; it is destroyed and then replaced by bone.
During all the years of growth there is constant remodelling with destruction (by osteoclasts) and replacement (by osteoblasts), whether the original development was intramembranous or endochondral. Similarly endochondral ossification, subperiosteal ossification and remodelling occurs in the callus of fracture sites.
The site where bone first forms is the primary centre of ossification, and in long bones is in the middle of the shaft (diaphysis), the centre first appearing about the eighth week of intrauterine life. The ends of the bone (epiphyses) remain cartilaginous and only acquire secondary ossification centres much later, usually after birth. The growing end of the diaphysis is the metaphysis, and the adjacent epiphyseal cartilage is the epiphyseal plate. When ossification occurs across the epiphyseal plate, the diaphysis and epiphysis fuse and bone growth ceases. The more actively growing end of a bone starts to ossify earlier and is the last to fuse with the diaphysis.
In the metaphysis the terminal branches of the nutrient artery of the shaft are end arteries, subject to the pathological phenomena of embolism and infarction; hence osteomyelitis in the child most commonly involves the metaphysis. The cartilaginous epiphysis has, like all hyaline cartilage, no blood supply. As ossification of the cartilaginous epiphysis begins, branches from the periarticular vascular plexus penetrate to the ossification centre. They have no communication across the epiphyseal plate with the vessels of the shaft. Not until the epiphyseal plate ossifies, at cessation of growth, are vascular communications established. Now the metaphysis contains no end arteries and is not subject to infarction from embolism; therefore osteomyelitis no longer has any particular site of election in the bone.
Sesamoid bones
Sesame seed-like sesamoid bones are usually associated with certain tendons where they glide over an adjacent bone. They may be fibrous, cartilaginous or bony nodules, or a mixture of all three, and their presence is variable. The only constant examples are the patella, which is by far the largest, and the ones in the tendons of adductor pollicis, flexor pollicis brevis and flexor hallucis brevis. In the foot they can also occur in the peroneus longus tendon over the cuboid, the tibialis anterior tendon against the medial cuneiform and the tibialis posterior tendon opposite the head of the talus. A sesamoid bone in the lateral head of gastrocnemius (the fabella) is not associated with a tendon. The reasons for the presence of sesamoids are uncertain. Sometimes they appear to be concerned in altering the line of pull of a tendon (patella in the quadriceps tendon) or with helping to prevent friction (as in the peroneus longus tendon moving against the cuboid bone).
Joints
Union between bones can be in one of three ways: by fibrous tissue; by cartilage; or by synovial joints.
Fibrous joints occur where bones are separated only by connective tissue (Fig. 1.4A) and movement between them is negligible. Examples of fibrous joints are the sutures that unite the bones of the vault of the skull and the syndesmosis between the lower ends of the tibia and fibula.
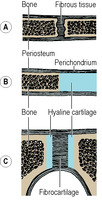
Cartilaginous joints are of two varieties, primary and secondary. A primary cartilaginous joint (synchondrosis) is one where bone and hyaline cartilage meet (Fig. 1.4B). Thus all epiphyses are primary cartilaginous joints, as are the junctions of ribs with their own costal cartilages. All primary cartilaginous joints are quite immobile and are very strong. The adjacent bone may fracture, but the bone–cartilage interface will not separate.
A secondary cartilaginous joint (symphysis) is a union between bones whose articular surfaces are covered with a thin lamina of hyaline cartilage (Fig. 1.4C). The hyaline laminae are united by fibrocartilage. There may be a cavity in the fibrocartilage, but it is never lined with synovial membrane and it contains only tissue fluid. Examples are the pubic symphysis and the joint of the sternal angle (between the manubrium and the body of the sternum). An intervertebral disc is part of a secondary cartilaginous joint, but here the cavity in the fibrocartilage contains a gel (p. 423).
A limited amount of movement is possible in secondary cartilaginous joints, depending on the amount of fibrous tissue within them. All symphyses occur in the midline of the body.
Typical synovial joints, which include all limb joints, are characterized by six features: the bone ends taking part are covered by hyaline cartilage and surrounded by a capsule enclosing a joint cavity, the capsule is reinforced externally or internally or both by ligaments, and lined internally by synovial membrane, and the joint is capable of varying degrees of movement. In atypical synovial joints the articular surfaces of bone are covered by fibrocartilage.
The synovial membrane lines the capsule and invests all non-articulating surfaces within the joint; it is attached round the articular margin of each bone. Cells of the membrane secrete a hyaluronic acid derivative which is responsible for the viscosity of synovial fluid, whose main function is lubrication. The viscosity varies, becoming thinner with rapid movement and thicker with slow. In normal joints the fluid is a mere film. The largest joint of all, the knee, only contains about 0.5mL.
The extent to which the cartilage-covered bone-ends make contact with one another varies with different positions of the joint. When the surfaces make the maximum possible amount of contact, the fully congruent joint is said to be close-packed (as in the knee joint in full extension). In this position the capsule and its reinforcing ligaments are at their tightest. When the surfaces are less congruent (as in the partly flexed knee), the joint is loose-packed and the capsule looser, at least in part.
Intra-articular fibrocartilages, discs or menisci, in which the fibrous element is predominant, are found in certain joints. They may be complete, dividing the joint cavity into two, or incomplete. They occur characteristically in joints in which the congruity between articular surfaces is low, e.g. the temporomandibular, sternoclavicular and knee joints.
Fatty pads are found in some synovial joints, occupying spaces where bony surfaces are incongruous. Covered in synovial membrane, they probably promote distribution of synovial fluid. The Haversian fat pad of the hip joint and the infrapatellar fold and alar folds of the knee joint are examples.
Mucous membranes
A mucous membrane is the lining of an internal body surface that communicates with the exterior directly or indirectly. This definition must not be taken to imply that all mucous membranes secrete mucus; many parts of the alimentary and respiratory tracts do, but most of the urinary tract does not. Mucous membranes consist of two and sometimes three elements: always an epithelium and an underlying connective tissue layer, the lamina propria, which in much of the alimentary tract contains a thin third component of smooth muscle, the muscularis mucosae. The whole mucous membrane, often called ‘mucosa’, usually lies on a further connective tissue layer, the submucous layer or submucosa. The epithelium of a mucous membrane varies according to the site and functional needs, e.g. stratified squamous in the mouth, columnar in the intestine, ciliated in the trachea.
Serous membranes
A serous membrane (serosa) is the lining of a closed body cavity—pericardial, pleural and peritoneal—and consists of connective tissue covered on the surface by a single layer of flattened mesothelial cells (derived from the mesoderm of the coelomic cavity). The part of the serosa that lines the wall of the cavity (the parietal layer of pericardium, pleura or peritoneum) is directly continuous with the same membrane that covers or envelops the mobile viscera within the cavity (the visceral layer). The peritoneal, pericardial and pleural cavities are potential slit-like spaces between the visceral layer and the parietal layer. The two layers slide readily on each other, lubricated by a film of tissue fluid. There are no glands to produce a lubricating secretion. The serous membranes are usually very adherent to the viscera. The parietal layer is attached to the wall of the containing cavity by loose areolar tissue and in most places can be stripped away easily.
The parietal layer of all serous membranes is supplied segmentally by spinal nerves. The visceral layer has an autonomic nerve supply.
Blood vessels
Blood vessels are of three types: capillaries; arteries; and veins.
Capillaries are the smallest vessels. Their walls consist only of flattened endothelial cells. Capillaries form an anastomotic network in most tissues. Certain structures, such as the cornea of the eye and hyaline cartilage, are devoid of capillaries.
Arteries conduct blood from the heart to the capillary bed, becoming progressively smaller, and as they do so, give way to arterioles which connect with the capillaries. Arterial walls have three layers. The tunica intima is very thin and comprises the endothelial lining, little collagenous connective tissue and an internal elastic lamina. Surrounding this layer is the tunica media consisting mainly of elastic connective tissue fibres and smooth muscle in varying amounts. The aorta and major arteries have a large proportion of elastic tissue which enables them to regain their original diameter after the expansion that follows cardiac contraction. Smaller arteries have less elastic tissue and more muscle. The tunica media of arterioles is almost entirely composed of smooth muscle. The outermost layer of the arterial wall is the tunica adventitia, which has an external elastic lamina surrounded by collagenous connective tissue.
Veins collect blood from the capillaries. They generally have a thinner wall and a larger diameter than their corresponding arteries. Veins have the same three layers in their walls as arteries, but a distinct internal elastic lamina is absent and there is much less muscle in the media. Peripheral limb veins are often double, as venae comitantes of their arteries. In the proximal parts of limbs venae comitantes unite into a single large vein. Many veins in the limbs and the neck have valves which prevent reflux of blood. These valves usually have two cup-shaped cusps formed by an infolding of the tunica intima. These cusps are apposed to the wall as long as the flow is towards the heart; when blood flow reverses, the valves close by assuming their cup-shaped form. On the cardiac side of a valve the vein wall is expanded to form a sinus. In general, there are no valves in the veins of the thorax and abdomen; testicular veins have valves.
Anastomoses between arteries are either actual or potential. In the former instance arteries meet end to end, such as the labial branches of the two facial arteries. A potential anastomosis is by terminal arterioles. Given sufficient time these arterioles can dilate to convey adequate blood, but with sudden occlusion of a main vessel the anastomosis is inadequate to immediately nourish the affected part, as in the case of the coronary arteries.
In many cases there is no precapillary anastomosis between adjacent arteries. Such vessels are end-arteries, and here interruption of arterial flow necessarily results in gangrene or infarction. Examples are found in the liver, spleen, kidney, lung, medullary branches of the central nervous system, the retina and the straight branches of the mesenteric arteries.
Arteriovenous anastomoses are short-circuiting channels between terminal arterioles and primary venules which occur in many parts of the body. They are plentiful in the skin, where they may have a role in temperature regulation.
Sinusoids are wide capillaries which have a fenestrated or discontinuous endothelium. They are numerous in the liver, spleen, adrenal medulla and bone marrow.
Blood vessels are innervated by efferent autonomic fibres which regulate the contraction of the smooth muscle in their walls. These nerves act on muscular arteries and especially on arterioles. Their main effect is vasoconstriction and increase in vascular tone, mediated by adrenergic sympathetic fibres. In some areas sympathetic cholinergic fibres inhibit muscle activity and cause vasodilatation. Circulating hormones and factors such as nitric oxide also act on vessel wall muscle. On account of the thickness of their walls, large vessels have their own vascular supply through a network of small vessels, the vasa vasorum.
Lymphatics
Not all the blood entering a part returns by way of veins; much of it becomes tissue fluid and returns by way of lymphatic vessels. Lymphatic capillaries are simple endothelial tubes. Larger collecting channels have walls similar to those of veins, but the specific tunics, or layers, are less distinct. They differ from veins in having many more valves. In general superficial lymphatics (i.e. in subcutaneous tissues) follow veins, while deep lymphatics follow arteries.
Clinical spread of disease (e.g. infection, neoplasm) by lymphatics does not necessarily follow strictly anatomical pathways. Lymph nodes may be bypassed by the disease process. If lymphatics become dilated by obstruction their valves may be separated and reversal of lymph flow can then occur. Lymphatics communicate with veins freely in many parts of the body; the termination of the thoracic duct in the neck may be ligated with impunity, for lymph finds its way satisfactorily into more peripheral venous channels.
Lymphoid tissue
The defence mechanisms of the body include phagocytosis, which is a non-specific engulfing process, and the immune response, which is a specific reaction to microorganisms and foreign proteins (antigens). The immune response may occur in two ways: (1) by the humoral antibody response, with production of antibodies which are protein molecules that circulate in the blood and attach themselves to the foreign protein so that the combination of antigen and antibody can be destroyed by phagocytosis; and (2) by the cell-mediated immune response, with the production of specific cells that circulate in the blood and destroy the antigen or stimulate its phagocytosis. Two types of lymphocyte produce these reactions: T cells are responsible for cell-mediated immunity and B cells for humoral antibody production. The B cells become transformed into plasma cells which produce the antibody molecules (the immunoglobulins: IgG; IgM; IgA; IgE; and IgD).
All lymphocytes arise from common stem cells in bone marrow (in the embryo from yolk sac, liver and spleen). Some of them circulate to and settle in the thymus, where they proliferate. After release into the bloodstream as T cells they colonize the spleen, lymph nodes and lymphoid follicles at other sites by passing through the postcapillary venules of those structures. Other stem cells become B cells and colonize lymphoid follicles without passing through the thymus. The T cells are so named because they depend on the thymus for their development; cell-mediated immunity thus depends on this organ. The B cells acquire their name from the bursa of Fabricius in birds, for it was in chickens that this organ (a diverticulum of the cloaca) was first found to be the source of humoral antibodies. The main types of T cell are cytotoxic T, helper T and regulatory T cells. B cells can either form plasma cells or become B memory cells.
The lymphoid organs consist of the thymus, lymph nodes and spleen. All are encapsulated and have an internal connective tissue framework to support the cellular elements. In all except the thymus the characteristic structural feature is the lymphoid nodule or follicle, which is typically a spherical collection of lymphocytes with a pale central area, the germinal centre. Unencapsulated lymphoid tissue occurs in mucosa-associated lymphoid tissue (MALT) in the mucosa and submucosa of the alimentary, respiratory and genitourinary tracts. Gut-associated lymphoid tissue (GALT) and bronchus-associated lymphoid tissue (BALT) are categories of MALT. Waldeyer’s peripharyngeal lymphoid ring of tonsils (palatine, lingual, nasopharyngeal and tubal) and Peyer’s patches in the ileum are areas of organized mucosa-associated lymphoid tissue (O-MALT). The overlying epithelium of these sites is able to sample antigens in the lumen and translocate them to the underlying lymphoid aggregation.
In the thymus the lymphocytes are not concentrated in rounded follicles but form a continuous dense band of tissue at the outer region or cortex of the lobules into which the organ is divided. The inner (paler) regions of the lobules form the medulla which has fewer lymphocytes and contains the characteristic thymic corpuscles (of Hassall); these are remnants of the epithelium of the third pharyngeal pouches from which the thymus developed.
In a typical lymph node the rounded follicles of lymphocytes are concentrated at the periphery (cortex). Lymphocytes, not collected into follicles, are also present in the paracortical areas and medullary region. B lymphocytes are found in the follicles and medulla; T lymphocytes in the paracortical areas and in the cortex between follicles. Several afferent lymph vessels enter through the capsule of the node and open into the subcapsular sinus. From here radial cortical sinuses drain to medullary sinuses which are confluent with the efferent vessel draining the node at the hilum, where blood vessels enter and leave. The thymus, spleen and the O-MALT aggregations, such as the tonsils, do not have afferent lymphatics.
The (palatine and pharyngeal) tonsils possess lymphoid follicles similar to those of lymph nodes, but while the nodes have a capsule of connective tissue the tonsils have, on their inner surfaces, a covering of mucous membrane that dips down deeply to form the tonsillar crypts.
The lymphoid follicles of the spleen are found in its white pulp, which is scattered in the red pulp that constitutes most of the substance of the spleen and contains large numbers of venous sinuses. In the white pulp T lymphocytes form periarteriolar sheaths. The sheaths are enlarged in places by lymphoid follicles with B lymphocytes in the germinal centres. These follicles are visible to the naked eye on the cut surface of the spleen as whitish nodules up to 1mm in diameter.
Apart from lymphocytes, all lymphoid organs and organized lymphoid tissue contain macrophages, which are part of the mononuclear phagocyte system of the body.
Part two. Nervous system
The nervous system is divided into the central nervous system, which consists of the brain and spinal cord, and the peripheral nervous system composed of cranial and spinal nerves and their associated ganglia. The central and peripheral parts each have somatic and autonomic components; the somatic are concerned with the innervation of skeletal muscle (along efferent pathways) and the transmission of sensory information (along afferent pathways), and the autonomic are concerned with the control of cardiac muscle, smooth muscle and glands (also involving efferent and afferent pathways). The term autonomic nervous system is applied collectively to all autonomic components.
Neurons and nerves
The structural and functional unit of the nervous system is the nerve cell or neuron. It consists of a cell body containing the nucleus, and a variable number of processes commonly called nerve fibres. A single cytoplasmic process, the axon (often very long), conducts nerve impulses away from the cell body, and may give off many collaterals and terminal branches to many different target cells. Other multiple cytoplasmic processes, the dendrites (usually very short), expand the surface area of the cell body for the reception of stimuli.
Pathways are established in the nervous system by communications between neurons at synapses, which are sites on the cell body or its processes where chemical transmitters enable nerve impulses to be handed on from one neuron to another. Transmission between neurons and cells outside the nervous system, for example muscle cells (neuromuscular junctions), is also effected by neuro-transmitters. The small number of ‘classic’ transmitters such as acetylcholine and noradrenaline (norepinephrine) has been vastly supplemented in recent years by many substances. These include monoamines, amino acids, nitric oxide and neuropeptides.
Cell bodies with similar function show a great tendency to group themselves together, forming nuclei within the central nervous system and ganglia outside it. Similarly processes from such aggregations of cell bodies tend to run together in bundles, forming tracts within the central nervous system and nerves outside the brain and spinal cord.
Apart from neurons the nervous system contains other cells collectively known as neuroglial cells (neuroglia or glia), which have supporting and other functions but which do not have the property of excitability or conductivity possessed by neurons. The main types of neuroglial cell are astrocytes and oligodendrocytes, which like neurons are developed from ectoderm of the neural tube. A third type of neuroglial cell is the microglial cell (microglia) which is the phagocytic cell of the nervous system, corresponding to the macrophage of connective tissue, and is derived from mesoderm.
Nerve fibres may be myelinated or unmyelinated. In the central nervous system myelin is formed by oligo-dendrocytes, and in peripheral nerves by Schwann cells (neurolemmocytes). In myelinated fibres, the regions where longitudinally adjacent Schwann cells or oligodendrocyte processes join one another are the nodes (of Ranvier). The white matter of the nervous system is essentially a mass of nerve fibres and is so called because of the general pale appearance imparted by the fatty myelin, in contrast to grey matter which is darker and consists essentially of cell bodies.
Peripheral nerve fibres have been classified in relation to their conduction velocity, which is generally proportional to size, and function:
• Group A—Up to 20 μm diameter, subdivided into:
α:12–20 μm. Motor and proprioception (Ia and Ib)
β:5–12 μm. Touch, pressure and proprioception (II)
γ:5–12 μm. Fusimotor to muscle spindles (II)
δ:1–15 μm. Touch, pain and temperature (III)
• Group B—Up to 3 μm diameter. Myelinated. Preganglionic autonomic
• Group C—Up to 2 μm diameter. Unmyelinated. Postganglionic autonomic, and touch and pain (IV).
The widest fibres tend to conduct most rapidly. Unfortunately, as can be seen from the above, it is not possible to make a precise prediction of function from mere size. Thus the largest myelinated fibres may be motor or proprioceptive and the smallest, whether myelinated or unmyelinated, are autonomic or sensory.
Spinal nerves
There are 31 pairs of spinal nerves: 8 cervical, 12 thoracic, 5 lumbar, 5 sacral and 1 coccygeal. Each spinal nerve is formed by the union of an anterior (ventral) and a posterior (dorsal) root which are attached to the side of the spinal cord by little rootlets. The union takes place within the intervertebral foramen through which the nerve emerges immediately distal to the swelling on the posterior root, the posterior root ganglion; most of these are also within the foramen. The anterior root of every spinal nerve contains motor (efferent) fibres for skeletal muscle; those from T1 to L2 inclusive and from S2 to S4 also contain autonomic fibres. The anterior root also contains a small number of unmyelinated afferent pain fibres which have ‘doubled back’ from their cells of origin in the posterior root ganglion to enter the spinal cord by the anterior root instead of by the posterior root. The posterior root of every nerve contains sensory (afferent) fibres whose cell bodies are in the posterior root ganglion. These are unipolar neurons, having a single process that bifurcates to pass to peripheral receptors and the central nervous system. Unlike in autonomic ganglia there are no synapses in posterior root ganglia.
Immediately after its formation the mixed spinal nerve divides into a larger anterior and a smaller posterior ramus. The great nerve plexuses—cervical, brachial, lumbar and sacral—are formed from anterior rami; posterior rami do not form plexuses.
Connective tissue binds the fibres of spinal nerves together to form the single nerve. Delicate loose connective tissue, the endoneurium, lies between individual fibres. Rounded bundles of fibres, or fascicles, are surrounded by the perineurium, a condensed layer of collagenous connective tissue. Fascicles are bound together into a single nerve by a layer of loose but thicker connective tissue, the epineurium. In the largest nerve, the sciatic, only about 20% of the cross-sectional area is nerve, so 80% is connective tissue, but in smaller nerves the amount of neural tissue is proportionally greater. The larger nerves have their own nerves, the nervi nervorum, in their connective tissue coverings.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


