CHAPTER 2 Integrating cells into tissues
Cells evolved as single, free-living organisms, but natural selection favoured more complex communities of cells, multicellular organisms, where groups of cells specialize during development to carry out specific functions for the body as a whole. This allowed the emergence of larger organisms with greater control over their internal environment and to the evolution of highly specialized organic structures such as the brain. The human body contains more than 200 different cell types, sharing the same genome but with different patterns of gene expression.
There is molecular evidence that this structure-based scheme of classification has validity. Thus the intermediate filament proteins (p. 15) characteristic of all epithelia are keratins; those of connective tissue are vimentins; those of muscle are desmins; and those of nervous tissue are neurofilament and glial fibrillary acidic proteins. However, cells such as myofibroblasts, neuroepithelial sensory receptors and ependymal cells of the central nervous system have features of more than one tissue type. Despite its anomalies, the scheme is useful for descriptive purposes and widely used, and will be adopted here.
In this section, two of the major tissue categories, epithelia and general connective and supporting tissues, will be described. Specialized skeletal connective tissues, i.e. cartilage and bone, together with skeletal muscle, are described in detail in Chapter 5 as part of the musculoskeletal system overview. Smooth muscle and cardiac muscle are described in Chapter 6. Nervous system tissues are described in Chapter 3. Specialized defensive cells, which also form a migrant population within the general connective tissues, are considered in more detail in Chapter 4, with blood, lymphoid tissues and haemopoiesis.
EPITHELIA
The term epithelium is applied to the layer or layers of cells that cover the body surfaces or line the body cavities that open on to it. Developmentally, epithelia are derived from all three layers of the early embryo (p. 198). The ectoderm gives rise to the epidermis, glandular tissue of the breast, cornea and the junctional zones of the buccal cavity and anal canal. The endoderm forms the epithelial lining of the alimentary canal and its glands, most of the respiratory tract and the distal parts of the urogenital tract. Mesodermal derivatives include the epithelia of the kidney, the suprarenal (adrenal) cortex and endocrine cells of the ovary and testis. These endocrine cells are atypical epithelia in that they differentiate from embryonic mesenchyme and, in common with endocrine cells in general, they lack a free surface that communicates with the exterior. This atypical category also includes endothelia that line blood vessels and lymphatics (p. 135), and the epithelium-like cell layers of mesodermal (and mesenchymal) origin that line internal cavities of the body and are usually classified separately as mesothelia: they line the pericardial, pleural and peritoneal cavities.
Epithelia function generally as selective barriers that facilitate, or inhibit, the passage of substances across the surfaces they cover. In addition, they may: protect underlying tissues against dehydration, chemical or mechanical damage; synthesize and secrete products into the spaces that they line; function as sensory surfaces. In this respect, many features of nervous tissue can be regarded as those of a modified epithelium and the two tissue types share an origin (p. 186) in embryonic ectoderm.
Epithelia (Fig. 2.1) are predominantly cellular and the little extracellular material they possess is limited to the basal lamina. Intercellular junctions, which are usually numerous, maintain the mechanical cohesiveness of the epithelial sheet and contribute to its barrier functions. A series of three intercellular junctions forms a typical epithelial junctional complex: in sequence from the apical surface, this consists of a tight junctional zone, an adherent (intermediate) junctional zone and a region of discrete desmosome junctions (p. 6). Epithelial cell shape is most usually polygonal and partly determined by cytoplasmic features such as secretory granules. The basal surface of an epithelium lies in contact with a thin layer of filamentous protein and proteoglycan termed the basal lamina, which is synthesized predominantly by the epithelial cells. The basal lamina is described on p. 33.
Epithelia can usually regenerate when injured. Indeed, many epithelia continuously replace their cells to offset cell loss caused by mechanical abrasion (reviewed in Blanpain et al 2007). Blood vessels do not penetrate typical epithelia and so cells receive their nutrition by diffusion from capillaries of neighbouring connective tissues. This arrangement limits the maximum thickness of living epithelial cell layers. Epithelia, together with their supporting connective tissue, can often be removed surgically as one layer, which is collectively known as a membrane. Where the surface of a membrane is moistened by mucous glands it is called a mucous membrane or mucosa (p. 39), whereas a similar layer of connective tissue covered by mesothelium is called a serous membrane or serosa (p. 39).
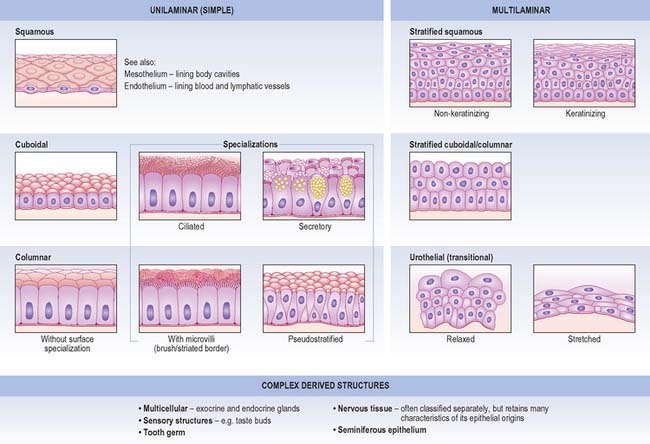
CLASSIFICATION
Unilaminar (simple) epithelia
Squamous epithelium
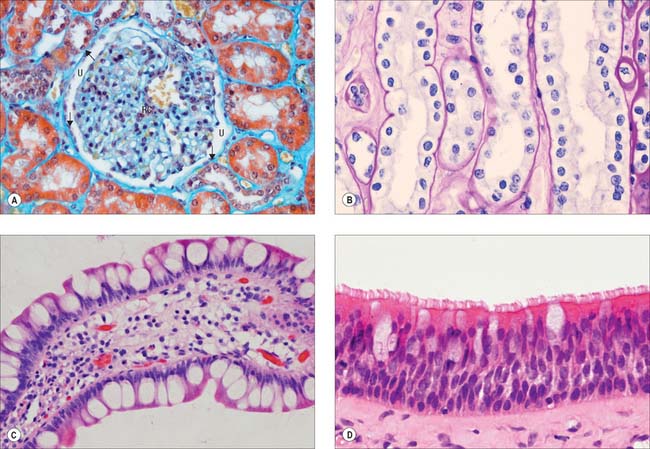
Simple squamous epithelium is composed of flattened, tightly apposed, polygonal cells (squames). This type of epithelium is described as tessellated when the cells have complex, interlocking borders rather than straight boundaries. The cytoplasm may in places be only 0.1 μm thick and the nucleus usually bulges into the overlying space (Fig. 2.2A). These cells line the alveoli of the lungs, where their surface area is huge and cytoplasmic volume correspondingly large, and they form the outer capsular wall of renal corpuscles, the thin segments of the renal tubules and various parts of the inner ear. Because it is so thin, simple squamous epithelium allows rapid diffusion of gases and water across its surface; it may also engage in active transport, as indicated by the presence of numerous endocytic vesicles in these cells. Tight junctions between adjacent cells ensure that materials pass primarily through cells, rather than between them.
Cuboidal and columnar epithelia
Cuboidal and columnar epithelia consist of regular rows of cylindrical cells (Fig. 2.2B, Fig. 2.2C). Cuboidal cells are approximately square in vertical section, whereas columnar cells are taller than their diameter, and both are polygonal when sectioned horizontally. Commonly, microvilli (Ch. 1) are found on their free surfaces, which considerably increases the absorptive area, e.g. in the epithelia of the small intestine (columnar cells with a striated border of very regular microvilli), the gallbladder (columnar cells with a brush border) and proximal convoluted tubules of the kidney (large cuboidal to low columnar cells with brush borders).
Ciliated columnar epithelium lines most of the respiratory tract, except for the lower pharynx and vocal folds, and it is pseudostratified (Fig. 2.2D) as far as the larger bronchioles; some of the tympanic cavity and auditory tube; the uterine tube; the efferent ductules of the testis. Submucosal mucous glands and mucosal goblet cells secrete mucus on to the luminal surface of much of the respiratory tract and cilia sweep a layer of mucus and trapped dust particles etc., from the lung towards the pharynx in the mucociliary rejection current, which clears the respiratory passages of inhaled particles. Cilia in the uterine tube assist the passage of oocytes and fertilized ova to the uterus (Ch. 77).
Some columnar cells are specialized for secretion and aggregates of such cells may be described as glandular tissue. Their apical domains (p. 4) typically contain mucus- or protein-filled (zymogen) vesicles, e.g. mucin-secreting and chief cells of the gastric epithelium. Where mucous cells lie among non-secretory cells, e.g. in the intestinal epithelium, their apical cytoplasm and its secretory contents often expand to produce a characteristic cell shape, and they are known as goblet cells (Fig. 2.2D). For further details of glandular tissue, see page 31 and for the characteristics of mucus, see page 39.
Pseudostratified epithelium
Pseudostratified epithelium is a single-layered (simple) columnar epithelium in which nuclei lie at different levels in a vertical section (Fig. 2.2D). All cells are in contact with the basal lamina throughout their lifespan, but not all cells extend through the entire thickness of the epithelium. Some constitute an immature basal cell layer of smaller cells, which are often mitotic and able to replace damaged mature cells. Migrating lymphocytes and mast cells within columnar epithelia may also give a similar, pseudostratified appearance because their nuclei are found at different depths. Much of the ciliated lining of the respiratory tract is of the pseudostratified type, and so is the sensory epithelium of the olfactory area.
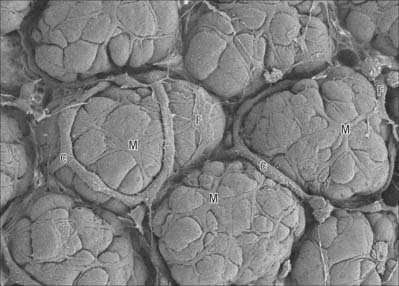
Myoepithelial cells
Myoepithelial cells, which are also sometimes termed basket cells, are fusiform or stellate in shape (Fig. 2.3), contain actin and myosin filaments, and contract when stimulated by nervous or endocrine signals. They surround the secretory portions and ducts of some glands, e.g. mammary, lacrimal, salivary and sweat glands, and lie between the basal lamina and the glandular or ductal epithelium. Their contraction assists the initial flow of secretion into larger conduits. Myoepithelial cells are ultrastructurally similar to smooth muscle cells in the arrangement of their actin and myosin, but differ from them because they originate, like the glandular cells, from embryonic ectoderm or endoderm. They can be identified immunohistochemically on the basis of the co-localization of myofilament proteins (which signify their contractile function), and keratin intermediate filaments (which accords with their epithelial lineage).
Multilaminar (stratified) epithelia
Stratified squamous epithelia
Stratified squamous epithelia are multilayered tissues in which the formation, maturation and loss of cells is continuous, although the rates of these processes can change, e.g. after injury. New cells are formed in the most basal layers by the mitotic division (p. 21) of stem cells and transit (or transient) amplifying cells. The daughter cells move more superficially, changing gradually from a cuboidal shape to a more flattened form and are eventually shed from the surface as a highly flattened squame. Typically, the cells are held together by numerous desmosomes to form strong, contiguous cellular sheets that provide protection to the underlying tissues against mechanical, microbial and chemical damage. Stratified squamous epithelia may be broadly subdivided into keratinized and non-keratinized types.
Keratinized epithelium
Keratinized epithelium (Fig. 2.4A) is found at surfaces that are subject to drying or mechanical stresses, or are exposed to high levels of abrasion. These include the entire epidermis and the mucocutaneous junctions of the lips, nostrils, distal anal canal, outer surface of the tympanic membrane and parts of the oral lining (gingivae, hard palate and filiform papillae on the anterior part of the dorsal surface of the tongue). Their cells, keratinocytes, are described in more detail on page 146. A distinguishing feature of keratinized epithelia is that cells of the superficial layer, the stratum corneum, are anucleate, dead, flattened squames that eventually flake off from the surface. In addition, the tough keratin intermediate filaments become firmly embedded in a matrix protein. This unusual combination of strongly coherent layers of living cells and more superficial strata made of plates of inert, mechanically robust protein complexes, interleaved with water-resistant lipid, makes this type of epithelium an efficient barrier against different types of injury, microbial invasion and water loss.
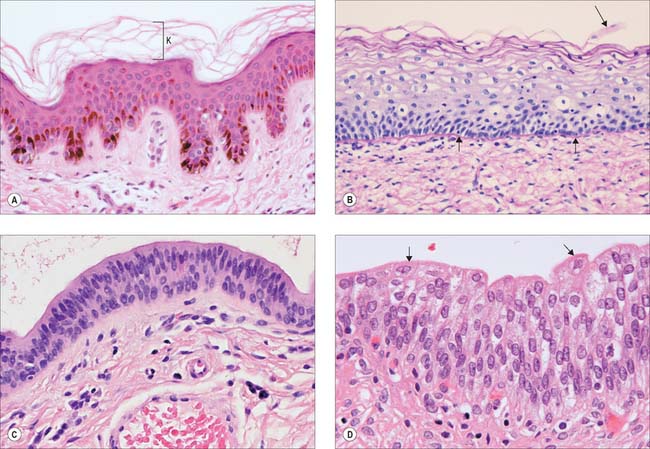
Non-keratinized epithelium
Non-keratinized epithelium is present at surfaces that are subject to abrasion but protected from drying (Fig. 2.4B). These include: the buccal cavity (except for the areas noted above); oropharynx and laryngopharynx; oesophagus; part of the anal canal; vagina; distal uterine cervix; distal urethra; cornea; inner surfaces of the eyelids; the vestibule of the nasal cavities. Cells go through the same transitions in general shape as are seen in the keratinized type, but they do not fill completely with keratin or secrete glycolipid, and they retain their nuclei until they desquamate at the surface. In sites where considerable abrasion occurs, e.g. parts of the buccal cavity, the epithelium is thicker and its most superficial cells may partly keratinize, so that it is referred to as parakeratinized, in contrast to the orthokeratinized state of fully keratinized epithelium. Diets deficient in vitamin A may induce keratinization of such epithelia, and excessive doses may lead to its transformation into mucus-secreting epithelium.
Stratified cuboidal and columnar epithelia
Two or more layers of cuboidal or low columnar cells (Fig. 2.4C) are typical of the walls of the larger ducts of some exocrine glands, e.g. the pancreas, salivary glands and the ducts of sweat glands and they presumably provide more strength than a single layer. Parts of the male urethra are also lined by stratified columnar epithelium. The layers are not continually replaced by basal mitoses and there is no progression of form from base to surface, but they can repair themselves if damaged.
Urothelium (urinary or transitional epithelium)
Urothelium (Fig. 2.4D) is a specialized epithelium that lines much of the urinary tract and prevents its rather toxic contents from damaging surrounding structures. It extends from the ends of the collecting ducts of the kidneys, through the ureters (p. 1242) and bladder (p. 1250), to the proximal portion of the urethra. In males it covers the urethra as far as the ejaculatory ducts, then becomes intermittent and is finally replaced by stratified columnar epithelium in the membranous urethra. In females it extends as far as the urogenital membrane. During development, part of it is derived from mesoderm and part from ectoderm and endoderm.
The epithelium appears to be four to six cells thick, and lines organs that undergo considerable distension and contraction. It can therefore stretch greatly without losing its integrity. In stretching, the cells become flattened, without altering their positions relative to each other, as they are firmly connected by numerous desmosomes. However, the urothelium appears to be reduced to two or three cells thick. The epithelium is called transitional because of the apparent transition between a stratified cuboidal epithelium and a stratified squamous epithelium, which occurs as it is stretched to accommodate urine, particularly in the bladder. The basal cells are basophilic, with many ribosomes, uninucleate (diploid), and are cuboidal when relaxed. More apically, they form large binucleate, or, more often, polyploid uninucleate cells. The surface cells are largest and may even be octaploid: in the relaxed state, they typically bulge into the lumen as dome-shaped cells with a thickened, eosinophilic glycocalyx or cell coat (p. 4). Their luminal surfaces are covered by a specialized plasma membrane in which plaques of intramembranous glycoprotein particles are embedded. These plaques stiffen the membrane. When the epithelium is in the relaxed state, and the surface area of the cells is reduced, the plaques are partially internalized by the hinge-like action of the more flexible interplaque membrane regions. They re-emerge onto the surface when it is stretched.
Seminiferous epithelium
Seminiferous epithelium is a highly specialized, complex stratified epithelium. It consists of a heterogeneous population of cells that form the lineage of the spermatozoa (spermatogonia, spermatocytes, spermatids), together with supporting cells (Sertoli cells). It is described in detail in Chapter 76 (p. 1266).
GLANDS
Glands may be subdivided into exocrine glands and endocrine glands. Exocrine glands secrete, usually via a duct, onto surfaces that are continuous with the exterior of the body, including the alimentary tract, respiratory system, urinary and genital ducts and their derivatives, and the skin. Endocrine glands are ductless and secrete hormones directly into interstitial fluid and thence the circulatory system, which conveys them throughout the body to affect the activities of other cells. In addition to strictly epithelial glands, some tissues derived from the nervous system, including the suprarenal medulla (p. 1200) and neuro-hypophysis (p. 321), are neurosecretory.
Paracrine glandular cells are similar to endocrine cells, but their secretions diffuse locally to cellular targets in the immediate vicinity; many of these are classed as neuroendocrine cells since they secrete molecules used elsewhere in the nervous system as neurotransmitters or neuromodulators (Ch. 3). Modes of signalling by secretory cells are illustrated in Fig. 1.6.
EXOCRINE GLANDS
Types of secretory process
The mechanism of secretion varies considerably. If the secretions are initially packaged into membrane-bound vesicles, these are conveyed to the cell surface (p. 9), where they are discharged. In merocrine secretion, which is by far the most common secretory mechanism, vesicle membranes fuse with the plasma membrane to release their contents to the exterior (Fig. 2.5). Specialized transmembrane molecules in the secretory vesicle wall recognize marker proteins on the cytoplasmic side of the plasma membrane and bind to them. This initiates interactions with other proteins that cause the fusion of the two membranes and the consequent release of the vesicle contents. The stimulus for secretion varies with the type of cell, but often appears to involve a rise in intracellular calcium. Glands such as the simple sweat glands of the skin, where ions and water are actively transported from plasma as an exudate, were once classified as eccrine glands. They are now known to synthesize and secrete small amounts of protein by a merocrine mechanism, and are thus reclassified as merocrine glands.
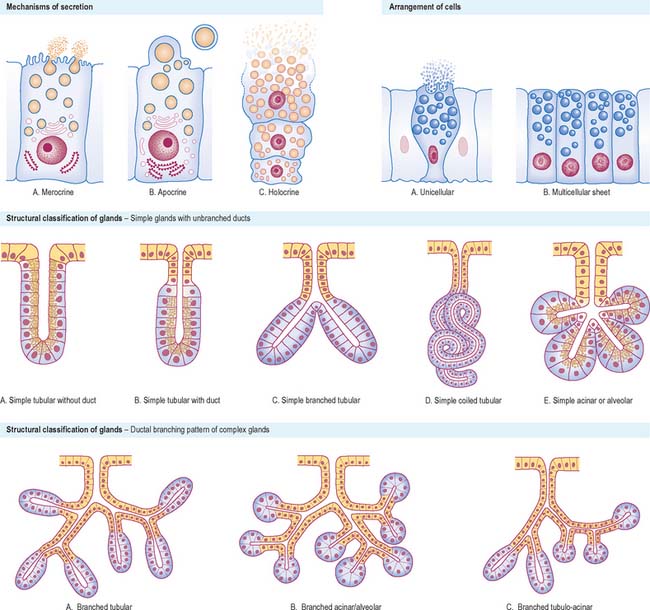
In apocrine glands, some of the apical cytoplasm is pinched off with the contained secretions, which are stored in the cell as membrane-free droplets (Fig. 2.5). The best understood example of this is the secretion of milk fat by mammary gland cells (p. 936), in which a small amount of cytoplasm is incorporated into the plasma membrane-bound lipid globule as it is released from the cell. Larger amounts of cytoplasm are included in secretions by specialized apocrine sweat glands in the axilla and anogenital regions of the body. In some tissues there is a combination of different types of secretion, e.g. mammary gland cells secrete milk fat by apocrine secretion and milk protein, casein, by merocrine secretion.
In holocrine glands (Fig. 2.5), e.g. sebaceous glands in the skin, the cells first fill with secretory products (lipid droplets or sebum, in this instance) and then the entire cell disintegrates to liberate the accumulated mass of secretion into the duct or, more usually, hair follicle.
Structural and functional classification
Exocrine glands are either unicellular or multicellular. The latter may be in the form of simple sheets of secretory cells, e.g. the lining of the stomach, or may be structurally more complex and invaginated to a variable degree. Such glands (Fig. 2.5) may be simple units or their connection to the surface may be branched. Simple unbranched tubular glands exist in the walls of many of the hollow viscera, e.g. the small intestine and uterus, whereas some simple glands have expanded, flask-like ends (acini or alveoli). Such glands may consist entirely of secretory cells, or may have a blind-ending secretory portion that leads through a non-secretory duct to the surface, in which case the ducts may modify the secretions as they pass along them.
ENDOCRINE GLANDS
Endocrine glands secrete directly into connective tissue interstitial fluid and the circulation. Their cells are grouped around beds of capillaries or sinusoids (p. 133) which typically are lined by fenestrated endothelia (Ch. 6
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


