Chapter Nine. General embryology
Embryology
Discoveries about the development of the human embryo are gained by research into the development of other species because of the problem of unethical human experimentation (The Human Fertilisation and Embryology Act 1990). There are surprising similarities in the basic machinery of development between species which involves cell division and differentiation, pattern formation, change in form and growth (Wolpert et al 2007).
The study of embryology enables us to understand the causes of congenital abnormalities, which are present in about 6% of live births, of which half are detected at birth and half during the first year of life. At least half of all conceptuses are malformed but are usually aborted spontaneously (Moore & Persaud 2008).
Gametogenesis
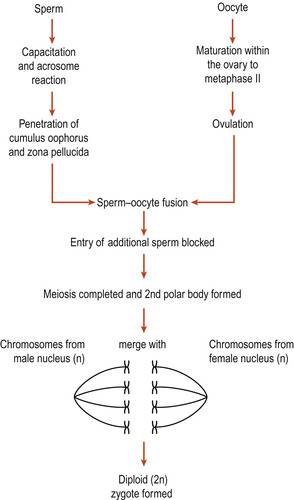
Following fertilisation of an ovum by a sperm the resulting zygote regains its full complement of 46 chromosomes. Gametogenesis is the formation of the sperm and ova. The primary spermatocytes and primary oocytes are diploid cells, having 46 chromosomes. Meiosis results in reduction of chromosomes to the haploid 23 found in the mature sperm and ovum (Fig. 9.1). Also independent assortment of maternal and paternal chromosomes amongst the gametes and crossing over between homologues of segments of the maternal and paternal chromosomes occurs (Ch. 3). This recombination of genetic material ensures that each gamete is a mixture of maternal and paternal genes as is the new zygote.
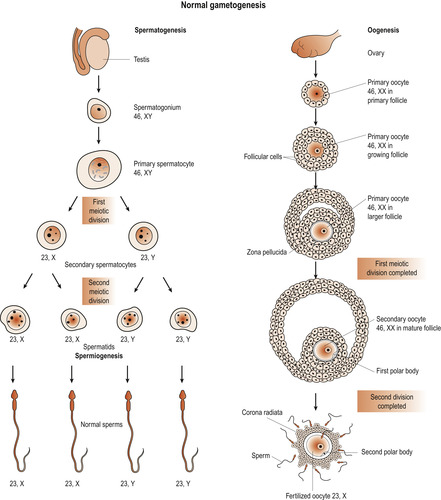 |
| Figure 9.1 Normal gametogenesis. (Reproduced with permission from Moore 1989.) |
Oogenesis
Primary oocytes are present in a female ovary before birth. Oogenesis or the process of transforming oocytes into ova begins before a woman’s birth but is not completed until after puberty. By the time a girl is born her primary oocytes have undergone the prophase of meiosis. Just before ovulation a surge of follicle-stimulating hormone (FSH) and luteinising hormone (LH) results in maturation of the ovum (Fig. 9.2) and the first meiotic division is completed (Fig. 9.3).
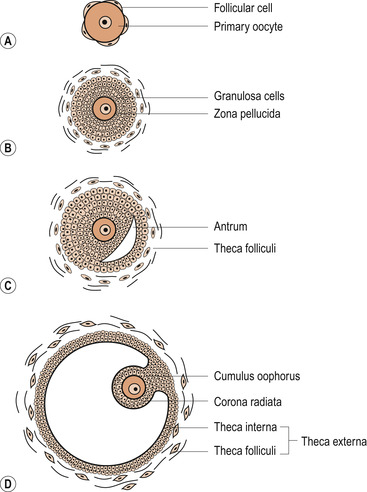 |
| Figure 9.2 The stages of development in the follicle: (A) primordial follicle, (B) primary follicle, (C) secondary follicle, (D) Graafian follicle. (From Hinchliff S M, Montague S E 1990, with permission.) |
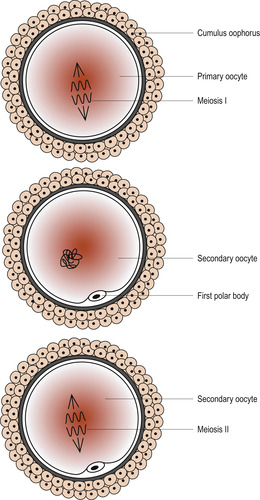 |
| Figure 9.3 Events within the zona pellucida prior to ovulation. (From Fitzgerald M J T, Fitzgerald M 1994, with permission.) |
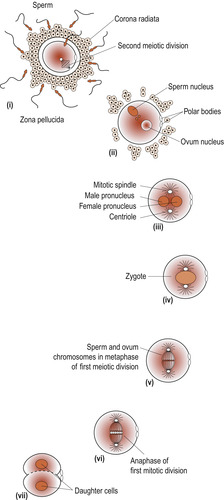
The process results in the formation of a secondary oocyte which receives most of the cytoplasm and a non-functional cell called the first polar body. The secondary oocyte receives 23 chromosomes, including an X chromosome, and the first polar body receives the other 23 chromosomes. At ovulation the secondary oocyte begins the second meiotic division but becomes arrested in metaphase (Fig. 9.4). If penetrated by a sperm, this division completes and one mature ovum with a second polar body results. These polar bodies degenerate (Schoenwolf et al 2008).
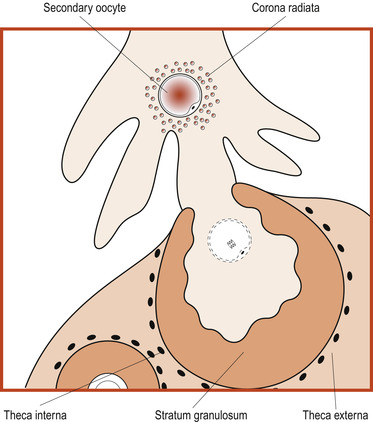 |
| Figure 9.4 Ovulation. (From Fitzgerald M J T, Fitzgerald M 1994, with permission.) |
Spermatogenesis
Spermatozoa are produced in the seminiferous tubules of the testes (Ch. 5). Primary spermatocytes begin to increase in number from puberty. In a functioning testis germ cells are present at various stages of development, all originating from spermatogonia which divide continuously by mitosis to ensure a constant supply of cells. Spermatogonia divide and grow to become primary spermatocytes, which are diploid cells with 46 chromosomes.
These undergo the first meiotic division to form two secondary spermatocytes which have only 23 chromosomes. Half receive an X chromosome and half a Y chromosome. Secondary meiosis results in four haploid spermatids found in close association with the Sertoli cells which provide nutrition and support. The spermatids are transformed into mature spermatozoa (Fig. 9.5).
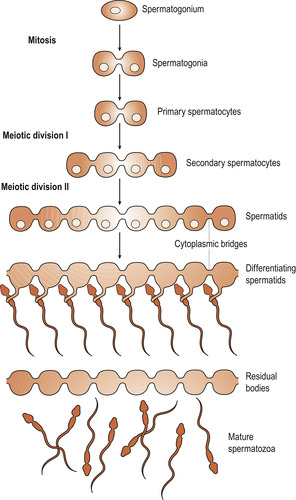 |
| Figure 9.5 The progeny of a single maturing spermatogonium remain connected to one another by cytoplasmic bridges throughout their differentiation into mature sperm. (Reproduced with permission from Alberts et al 1994.) |
As a sperm matures, excess protoplasm is lost and the nuclear chromatin condenses to become the head of the sperm. One centriole develops into the tail which is composed of a central filament of two microfibrils surrounded by a circle of nine fibrils. Mitochondria aggregate in the neck region and the Golgi apparatus helps to form the acrosome cap which develops over the head of the sperm and contains enzymes called hyaluronidases and proteases (Fig. 9.6).
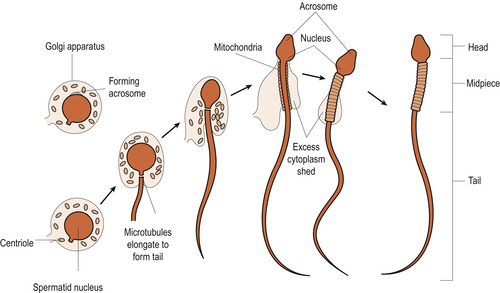 |
| Figure 9.6 Sperm formation. (Reproduced with permission from Chiras 1991.) |
Gamete size
The oocyte is a very large cell, just visible to the unaided eye and usually only one is released at ovulation. It contains all the material necessary for early embryonic growth and development. In sharp contrast, the mature sperm has lost most of its cytoplasm, is very small and millions are released at ejaculation. About 1000 sperm will reach the oocyte and these will all be needed to allow just one to enter to form a diploid zygote, the first cell of a unique human being.
Fertilisation
Capacitation
Freshly ejaculated sperm are unable to fertilise an ovum and have to undergo a process of maturation called capacitation. The sperm have to penetrate several physical barriers before entering the ovum. While travelling through the female genital tract, usually in the uterus or uterine tubes, glycoproteins are removed from the surface of the acrosome cap. When a capacitated sperm meets the corona radiata of the oocyte the acrosome develops perforations in it and the contents of the acrosomal vesicle are released. This is known as the acrosome reaction.
The acrosome reaction
Binding of a sperm of the oocyte’s zona pellucida is species-specific. Lytic (digestive) enzymes such as hyaluronidases are released around the oocyte and digest the first physical barrier of cumulus cells embedded in sticky hyaluronic acid. It takes enzymes from many sperm to allow one sperm to enter. These enzymes disperse the corona radiata follicular cells, allowing the head of one sperm to make contact with the zona pellucida. Other enzymes such as acrosin, which produce an opening in the zona pellucida, are released (Wolpert et al 2007). The sperm cell membrane and the sperm nucleus pass into the ovum (Fig. 9.7).
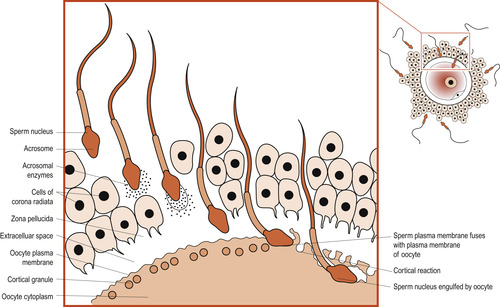 |
| Figure 9.7 Fertilisation and cortical reaction. (Reproduced with permission from Chiras 1991.) |
Blocks to polyspermy
Two mechanisms prevent polyspermy (the entry of multiple sperm) immediately following the entry of the first sperm, ensuring that the fertilised ovum only contains 46 chromosomes (Fig. 9.8).
• Fast block—The electrical resting potential of the oocyte plasma membrane is normally negatively charged at −70 millivolts (mV), at which sperm can readily fuse with the membrane. Immediately after sperm entry sodium channels open in the cell membrane and extra positively charged sodium ions are allowed into the cytoplasm of the ovum. This changes the charge to a positive +20 mV which prevents other sperm from entering. This is a brief reaction and the resting potential soon returns to −70 mV.
• Slow bock (cortical reaction)—The brief depolarisation has two other effects. First it allows subcortical granules lying just under the oocyte’s plasma membrane to rupture and release chemicals which bind water into the cytoplasm. The ovum swells and detaches any remaining sperm in contact with it. Secondly, the secondary oocytes are activated to complete the secondary meiotic division and expel the second polar body and the ovum is now mature. Its nucleus is now called the female pronucleus. Once the head of the sperm enters the cytoplasm of the ovum its head enlarges to form the male pronucleus. The male and female pronuclei fuse and paternal and maternal chromosomes intermingle.
 |
| Figure 9.8 Events in the female reproductive tract leading up to fertilisation. (From Hinchliff S M, Montague S E 1990, with permission.) |
Mitochondrial deoxyribonucleic acid (MtDNA)
The embryo
Terminology
The term conceptus refers to the products of fertilisation and comprises the embryo with its supporting tissues (adnexi). The developmental process is divided into discrete sections of time. The preimplantation period is the time between fertilisation and implantation and lasts about 6 days. The conceptus is called an embryo from implantation until the end of the 8th week after fertilisation when it becomes known as a fetus. The size of the embryo is expressed as the crown–rump length, from the crown of the head to the terminal part of the caudal end. In the fetus, the standing length from crown to heel is used.
General concepts used in embryology
How does one cell develop into millions with hundreds of variations? What instructions does the cell use and what processes carry out the instructions? There are three important concepts towards understanding embryology:
1. A programme of simple instructions can generate complex forms.
2. Most developmental processes depend upon interaction between genetic and environmental factors.
3. Each body system has its own developmental pattern.
Programming the embryo
The information to make an embryo is located in the DNA of the zygote. Although the sperm and ovum contribute 23 chromosomes each, it is the ovum that contributes the organelles and MtDNA. Regulatory genes control development by influencing where and when proteins are made. Developmental processes include (Wolpert et al 2007):
• Pattern formation.
• Differentiation.
• Morphogenesis.
• Development of the germ layers.
• Growth.
Pattern formation
Cells in the embryo seem to ‘know’ where and when to change shape and position and cell movements are part of the embryo’s developmental programme. Wolpert (1991) described this as a set of instructions not for describing the final form, but for creating shapes. Consider knitting: the pattern is the code or instruction manual but does not resemble the final article. The knitter must interpret the code and apply energy, order and control and a variety of stitches.
Cellular processes include:
• Somatic cell division where daughter cells receive identical genetic information.
• Cell differentiation to make up the different embryological tissues.
• Induction: cell interaction where one type of cell influences another.
• Migration of cells.
• Programmed cell death to remove redundant cells.
Early cell division—cleavage
Early in pregnancy growth and development are very similar in any human embryo. The exact days when embryonic features develop can be given. Days of development are counted from the moment of fertilisation. Within 24 h the large zygote undergoes mitosis and splits into smaller cells (Fig. 9.9). This is called cleavage and the daughter cells are called blastomeres. There is synthesis of new DNA but no increase in the amount of cytoplasm so the size of the blastomeres diminishes with each cleavage (Moore & Persaud 2008). The zygote continues its journey down the uterine tube and by the fourth day there are between 16 and 20 cells. Compaction occurs, probably because of cell surface adhesion glycoproteins. The embryo is now called a morula (Fig. 9.10). The cells are still totipotent and could contribute to any part of the embryo or even split to form identical fetuses.