OBJECTIVES
After studying this chapter, you should be able to:
List the major forms of motility in the gastrointestinal tract and their roles in digestion and excretion.
Distinguish between peristalsis and segmentation.
Explain the electrical basis of gastrointestinal contractions and the role of basic electrical activity in governing motility patterns.
Describe how gastrointestinal motility changes during fasting.
Understand how food is swallowed and transferred to the stomach.
Define the factors that govern gastric emptying and the abnormal response of vomiting.
Define how the motility patterns of the colon subserve its function to desiccate and evacuate the stool.
INTRODUCTION
The digestive and absorptive functions of the gastrointestinal system outlined in the previous chapter depend on a variety of mechanisms that soften the food, propel it through the length of the gastrointestinal tract (Table 27–1), and mix it with bile from the gallbladder and digestive enzymes secreted by the salivary glands and pancreas. Some of these mechanisms depend on intrinsic properties of the intestinal smooth muscle. Others involve the operation of reflexes involving the neurons intrinsic to the gut, reflexes involving the central nervous system (CNS), paracrine effects of chemical messengers, and gastrointestinal hormones.
GENERAL PATTERNS OF MOTILITY
Peristalsis is a reflex response that is initiated when the gut wall is stretched by the contents of the lumen, and it occurs in all parts of the gastrointestinal tract from the esophagus to the rectum. The stretch initiates a circular contraction behind the stimulus and an area of relaxation in front of it (Figure 27–1). The wave of contraction then moves in an oral-to-caudal direction, propelling the contents of the lumen forward at rates that vary from 2 to 25 cm/s. Peristaltic activity can be increased or decreased by the autonomic input to the gut, but its occurrence is independent of extrinsic innervation. Indeed, progression of the contents is not blocked by removal and resuture of a segment of intestine in its original position and is blocked only if the segment is reversed before it is sewn back into place. Peristalsis is an excellent example of the integrated activity of the enteric nervous system. It appears that local stretch releases serotonin, which activates sensory neurons that activate the myenteric plexus. Cholinergic neurons passing in a retrograde direction in this plexus activate neurons that release substance P and acetylcholine, causing smooth muscle contraction behind the bolus. At the same time, cholinergic neurons passing in an anterograde direction activate neurons that secrete NO and vasoactive intestinal polypeptide (VIP), producing the relaxation ahead of the stimulus.
FIGURE 27–1
Patterns of gastrointestinal motility and propulsion. An isolated contraction moves contents orally and aborally. Segmentation mixes contents over a short stretch of intestine, as indicated by the time sequence from left to right. In the diagram on the left, the vertical arrows indicate the sites of subsequent contraction. Peristalsis involves both contraction and relaxation, and moves contents aborally.
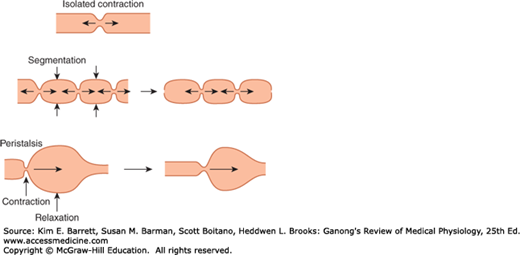
When the meal is present, the enteric nervous system promotes a motility pattern that is related to peristalsis but is designed to retard the movement of the intestinal contents along the length of the intestinal tract to provide time for digestion and absorption (Figure 27–1). This motility pattern is known as segmentation, and it provides for ample mixing of the intestinal contents (known as chyme) with the digestive juices. A segment of bowel contracts at both ends, and then a second contraction occurs in the center of the segment to force the chyme both backward and forward. Unlike peristalsis, therefore, retrograde movement of the chyme occurs routinely in the setting of segmentation. This mixing pattern persists for as long as nutrients remain in the lumen to be absorbed. It presumably reflects programmed activity of the bowel dictated by the enteric nervous system, and can occur independent of central input, although the latter can modulate it.
Except in the esophagus and the proximal portion of the stomach, the smooth muscle of the gastrointestinal tract has spontaneous rhythmic fluctuations in membrane potential between about −65 and −45 mV. This basic electrical rhythm (BER) is initiated by the interstitial cells of Cajal, stellate mesenchymal pacemaker cells with smooth muscle-like features that send long multiply branched processes into the intestinal smooth muscle. In the stomach and the small intestine, these cells are located in the outer circular muscle layer near the myenteric plexus; in the colon, they are at the submucosal border of the circular muscle layer. In the stomach and small intestine, there is a descending gradient in pacemaker frequency, and as in the heart, the pacemaker with the highest frequency usually dominates.
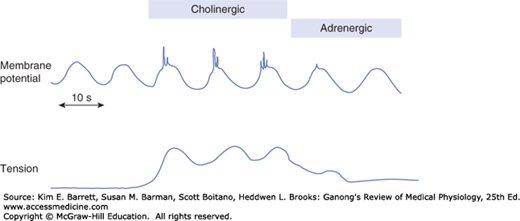
The BER itself rarely causes muscle contraction, but spike potentials superimposed on the most depolarizing portions of the BER waves do increase muscle tension (Figure 27–2). The depolarizing portion of each spike is due to Ca2+ influx, and the repolarizing portion is due to K+ efflux. Many polypeptides and neurotransmitters affect the BER. For example, acetylcholine increases the number of spikes and the tension of the smooth muscle, whereas epinephrine decreases the number of spikes and the tension. The rate of the BER is about 4/min in the stomach. It is about 12/min in the duodenum and falls to about 8/min in the distal ileum. In the colon, the BER rate rises from about 2/min at the cecum to about 6/min at the sigmoid. The function of the BER is to coordinate peristaltic and other motor activity, such as setting the rhythm of segmentation; contractions can occur only during the depolarizing part of the waves. After vagotomy or transection of the stomach wall, for example, peristalsis in the stomach becomes irregular and chaotic.
During fasting between periods of digestion, the pattern of electrical and motor activity in gastrointestinal smooth muscle becomes modified so that cycles of motor activity migrate from the stomach to the distal ileum. Each cycle, or migrating motor complex (MMC), starts with a quiescent period (phase I), continues with a period of irregular electrical and mechanical activity (phase II), and ends with a burst of regular activity (phase III) (Figure 27–3). The MMCs are initiated by motilin. The circulating level of this hormone increases at intervals of approximately 100 min in the interdigestive state, coordinated with the contractile phases of the MMC. The contractions migrate aborally at a rate of about 5 cm/min, and also occur at intervals of approximately 100 min. Gastric secretion, bile flow, and pancreatic secretion increase during each MMC. They likely serve to clear the stomach and small intestine of luminal contents in preparation for the next meal.
FIGURE 27–3
Migrating motor complexes (MMCs). The three phases include a quiescent phase (phase I); a phase consisting of small, irregular contractions that do not propagate (phase II); and a phase of regular activity lasting about 5 minutes (phase III), which sweeps along the length of the intestine. The entire cycle repeats every 90–100 minutes under fasting conditions. Note that the complexes are completely inhibited by a meal and resume 90–120 min later.
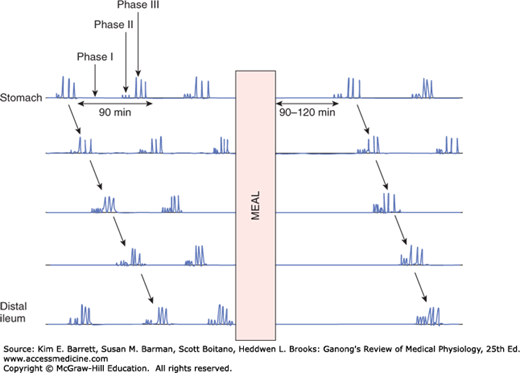
Conversely, when a meal is ingested, secretion of motilin is suppressed (ingestion of food suppresses motilin release via mechanisms that have not yet been elucidated), and the MMC is abolished, until digestion and absorption are complete. Instead, there is a return to peristalsis and the other forms of BER and spike potentials during this time. The antibiotic erythromycin binds to motilin receptors, and derivatives of this compound may be of value in treating patients in whom gastrointestinal motility is decreased.
SEGMENT-SPECIFIC PATTERNS OF MOTILITY
In the mouth, food is mixed with saliva and propelled into the esophagus. Peristaltic waves in the esophagus move the food into the stomach.
Chewing (mastication) breaks up large food particles and mixes the food with the secretions of the salivary glands. This wetting and homogenizing action aids swallowing and subsequent digestion. Large food particles can be digested, but they cause strong and often painful contractions of the esophageal musculature. Particles that are small tend to disperse in the absence of saliva and also make swallowing difficult because they do not form a bolus. The number of chews that is optimal depends on the food, but usually ranges from 20 to 25.
Edentulous patients are generally restricted to a soft diet and have considerable difficulty eating dry food.
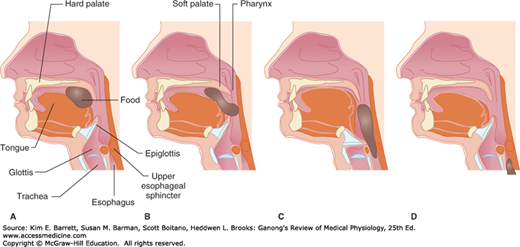
Swallowing (deglutition) is a reflex response that is triggered by afferent impulses in the trigeminal, glossopharyngeal, and vagus nerves (Figure 27–4). These impulses are integrated in the nucleus of the tractus solitarius and the nucleus ambiguus. The efferent fibers pass to the pharyngeal musculature and the tongue via the trigeminal, facial, and hypoglossal nerves. Swallowing is initiated by the voluntary action of collecting the oral contents on the tongue and propelling them backward into the pharynx. This starts a wave of involuntary contraction in the pharyngeal muscles that pushes the material into the esophagus. Inhibition of respiration and glottic closure are part of the reflex response. A peristaltic ring contraction of the esophageal muscle forms behind the material, which is then swept down the esophagus at a speed of approximately 4 cm/s. When humans are in an upright position, liquids and semisolid foods generally fall by gravity to the lower esophagus ahead of the peristaltic wave. However, if any food remains in the esophagus, it is cleared by a second wave of peristalsis that occurs by the mechanisms discussed above. It is therefore possible to swallow food while standing on one’s head.
FIGURE 27–4
Movement of food through the pharynx and upper esophagus during swallowing. A) The tongue pushes the food bolus to the back of the mouth. B) The soft palate elevates to prevent food from entering the nasal passages. C) The epiglottis covers the glottis to prevent food from entering the trachea and the upper esophageal sphincter relaxes. D) Food descends into the esophagus.
Unlike the rest of the esophagus, the musculature of the gastroesophageal junction (lower esophageal sphincter; LES) is tonically active but relaxes on swallowing. The tonic activity of the LES between meals prevents reflux of gastric contents into the esophagus. The LES is made up of three components (Figure 27–5)
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


