CHAPTER 51 Development of the limbs
Development of the limb depends on a series of complex cell–cell interactions that have been identified in experiments on amphibian, avian and reptilian species; the range demonstrates the remarkable conservation of developmental processes. Fate maps have revealed which structures in the limb arise from particular cells or groups of cells in the embryo and considerable progress has been made in identifying the genetic basis of limb development. The finding that the same molecules are involved in developing embryos of both model organisms and humans has promoted an increasing convergence between developmental biology and clinical genetics.
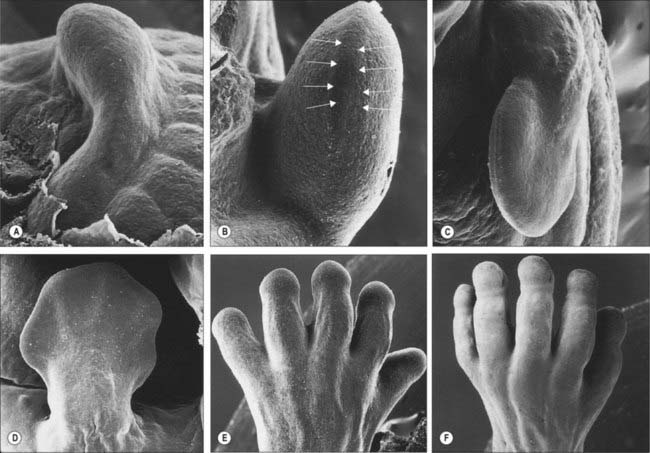
The limbs develop from somatopleuric mesenchyme in the lateral body wall. Regions of the somatopleuric mesenchyme at specific positions along the main body axis proliferate extensively to give rise to limb buds. These buds are the first visible signs of limb development and are rimmed by a longitudinal ridge of high columnar epithelial cells, the apical ectodermal ridge (Fig. 51.1).
AXES OF LIMBS
For descriptive, experimental and conceptual purposes, it has been necessary to define and name various ‘axes’, borders, surfaces and lines in relation to the developing limb bud (Fig. 51.2). An imaginary line from the centre of the elliptical base of the bud, through the centre of its mesenchymal core, to the centre of the apical ectodermal ridge, defines the proximodistal axis of the limb bud (previously known in descriptive embryology simply as the axis). Named in relation to the latter, the cranially placed limb border is the preaxial border, and the caudally placed limb border is the postaxial border. (In tetrapods and birds, the last two are termed anterior and posterior borders, respectively.) Any line that passes through the limb bud from preaxial to postaxial border, orthogonal to the proximodistal axis, constitutes a craniocaudal axis. The dorsal and ventral ectodermal surfaces thus clothe their respective aspects from preaxial to postaxial borders, and any line that passes from dorsal to ventral aspect, orthogonal to both proximodistal and craniocaudal axes, constitutes a dorsoventral axis. It should be noted here that the terms dorsal and ventral axial lines are to be used exclusively in relation to developing and definitive patterns of cutaneous innervation of the limbs and their associated levels of the trunk.
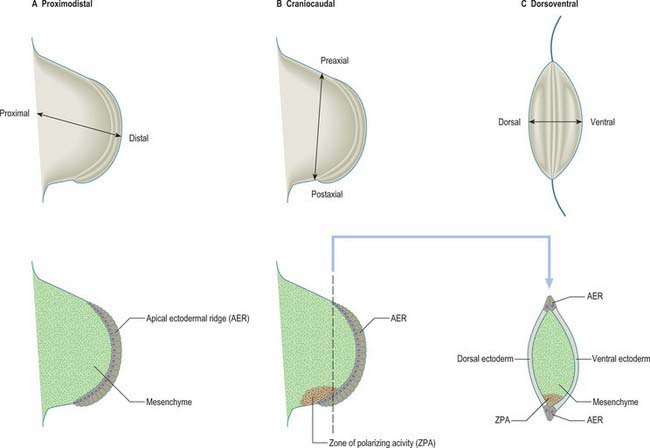
The three developmental axes (proximodistal, craniocaudal, and dorsoventral) can be identified in the developing limb bud by stage 13. Experiments in chick embryos have shown that development of structures in relation to each of these three principal axes seem to be specified by different mechanisms (Fig. 51.2). Outgrowth along the proximodistal axis is controlled by the apical ectodermal ridge and subjacent somatopleuric mesenchyme; outgrowth of the limb bud is accompanied by the sequential formation of limb structures along the proximodistal axis. The craniocaudal axis is controlled by a small population of mesenchymal cells on the postaxial border of the limb bud; this region of mesenchyme is termed the zone of polarizing activity or polarizing region. The dorsoventral axis of the limb appears to be controlled by the surface ectodermal covering of the limb bud.
APICAL ECTODERMAL RIDGE
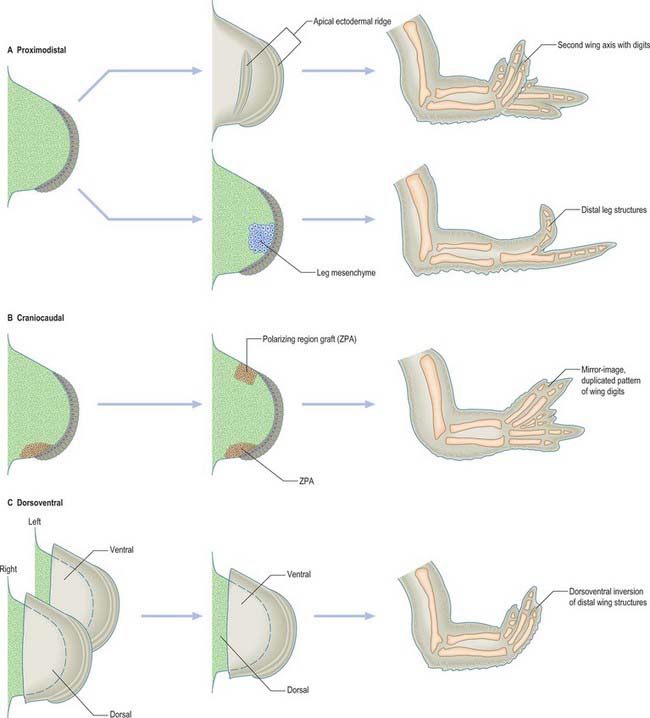
The fundamental epithelial–mesenchymal interactions seen in limb development are discussed in detail in Hinchcliffe & Johnson (1980) and the more recently discovered genetic basis of these interactions in Ferretti & Tickle (2006). The knowledge of limb embryology gained from studying chicks may be summarized as follows (Fig. 51.3). The apical ectodermal ridge provides the orientating influence for limb bud outgrowth. Its removal results in cessation of limb development, whereas grafting a second apical ectodermal ridge results in two axes of development, and duplication of distal structures (Fig. 51.3). There is evidence that the limb mesenchyme beneath the apical ectodermal ridge provides an apical ectodermal ridge maintenance factor that is essential to the function of the ridge. Thus, when leg bud cells are grafted to the tip of a wing bud beneath the apical ectodermal ridge, the ridge is maintained and bud outgrowth continues (it should be noted that the leg cells, even though placed in a wing bud, still form distal leg structures) (Fig. 51.3). In addition, the leg mesenchyme will pass information to the local ectoderm eliciting the appropriate epidermal development, which in this case is the formation of scales rather than feathers.
ZONE OF POLARIZING ACTIVITY
The zone of polarizing activity is a small region of somatopleuric mesenchyme on the postaxial border of the limb. When a second zone of polarizing activity is grafted beneath or adjacent to the apical ectodermal ridge at the preaxial border of a chick wing bud, duplication of distal limb structures occurs, generating a mirror-image pattern of six digits (Fig. 51.3). The digit closest to the zone of polarizing activity is always the most postaxial digit (in the chick wing this is digit 4), while more preaxial digits develop further away from the zone of polarizing activity. Thus signalling from the zone of polarizing activity appears to control both digit number and digit pattern. It has been suggested that digit number is controlled by regulation of the production of the apical ectodermal ridge maintenance factor, while digit pattern is regulated by production of a diffusible morphogen that acts in a concentration dependent fashion.



