CHAPTER 35 Development of the head and neck
Head development is distinct from that of the trunk, utilizing region-specific genes, signalling mechanisms and morphogenetic processes. This chapter will provide a description of head and neck development based primarily on data from studies on human embryos. Since this information is necessarily purely descriptive, it will be supplemented by observations from lineage and genetic studies in mouse embryos, where the results are likely to be valid for all mammals, including human. Human studies are taken from the following sources, which incorporate primary data by the authors as well as references to older primary source material: O’Rahilly and Müller (1987, 2001); Hinrichsen (1990). References to experimental studies in the mouse are provided in the text.
Evolution of the vertebrate head was made possible by the origin of a novel cell population, the neural crest. Neural crest cells have the potential to form connective and skeletal tissues in the head, and they make major contributions to the skull. In mammalian embryos, cranial neural crest cells emigrate from the edges of the still unfused cranial neural folds, unlike trunk neural crest cells and the cranial crest of other vertebrates, which begin migration only after neural tube closure. Figure 35.1 shows stylized views of human embryos at an early stage of neural crest migration (A) and at the end of the crest migration (B). N.B. These views do not show the neural crest cells themselves and not by a specific staining procedure.
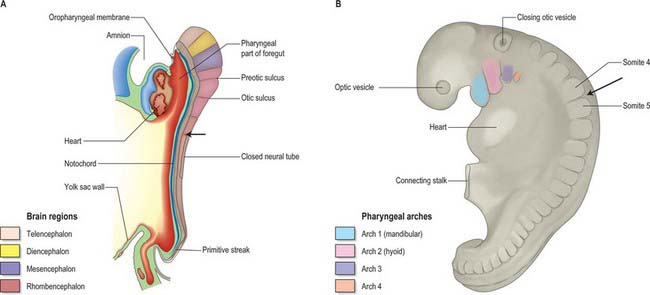
Fig. 35.1 Human embryos during early stages of head development. The occipital–cervical boundary is indicated by an arrow. A. Sagittal section of a stage 10 embryo in which neural tube closure has begun in the future occipito-cervical region. B. Lateral view of a stage 12 embryo; the occipital/cervical boundary between somites 4 and 5 is indicated by an arrow. (See also fig 24.2).
In the mouse, a lineage marker has enabled neural crest cells to be traced from the stage at which they leave the neural epithelium to their locations in adult tissues. These studies show three separate populations of neural crest cells migrating from the cranial neural folds which give rise to neuronal and non-neuronal progeny (Fig. 35.2). The first of these populations (trigeminal) originates from the diencephalic region of the forebrain, the midbrain, and prorhombomere A of the hindbrain, which subsequently divides to form the first two rhombomeres (Fig. 35.2A,B; see also Fig. 24.2). Those neural crest cells with a neuronal fate contribute to the trigeminal ganglion (Jiang et al 2002). The non-neuronal mesenchymal crest cells migrate extensively to surround the telencephalon and part of the diencephalon, forming the frontonasal mesenchymal populations; they also migrate lateral to the rhombencephalon to form the mesenchyme of the maxillary and mandibular regions of the first arch (Fig. 35.2C,D). The second (hyoid) population gives rise to the otic ganglion and then migrates from prorhombomere B (which forms rhombomeres 3 and 4), into the second pharyngeal arch. The third (vagal) population has a more extensive origin, from the neural folds caudal to the otocyst, i.e. prorhombomere C. These cells will contribute to the ganglia of the glossopharyngeal and vagal nerves, with the non-neuronal vagal crest cells migrating into pharyngeal arches 3, 4 and 6, some of them continuing into the heart to contribute to the division of the cardiac outflow tract. The neural crest also gives rise to the parasympathetic ganglia and parasympathetic postganglionic nerves in the head and neck. For a full account of the neuronal contribution of neural crest cells, see Chapter 24. In human embryos, histological methods have revealed equivalent cranial neural crest cell origins and migration routes to those of the mouse, except that no emigration from the diencephalon has been detected (O’Rahilly & Müller 2007).
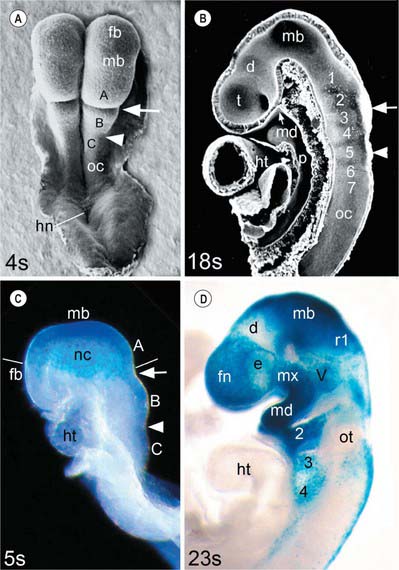
Fig. 35.2 Mouse embryo heads during the period of cranial neural crest cell migration. A and B, Scanning electron micrographs (A, dorsal view; B, median section) of embryos near the start (4 somite pairs) and end (18 somite pairs, interior view) of the period of neural crest cell migration. At the 4-somite stage the neural folds are convex and the hindbrain is divided into prorhombomeres A, B and C by the preotic (arrow) and otic (arrowhead) sulci; at the 18-somite stage the neural tube is closed and the hindbrain is divided into seven rhombomeres (numbered) and the unsegmented occipital region (oc). C and D, Lateral views of 5- and 23-somite stage embryos carrying a permanent marker for neural crest cells and the neural epithelium from which the trigeminal crest cells originate (blue stain). Neural crest cells (nc) migrate as a veil-like mass beneath the surface ectoderm, originating as a continuous group from prorhombomere A, the midbrain and the caudal diencephalic region of the forebrain, into the frontonasal region (fn) and the first arch (lines indicate the level of the migrating edge). After migration, frontonasal neural crest cells (fn) cover the telencephalon and rostral diencephalon, surrounding the eye (e); maxillary (mx) and mandibular (md) neural crest populates the first pharyngeal arch; the trigeminal ganglion primordium (V) is also present. Crest cells from prorhombomere B have populated the hyoid arch and the primordium of ganglion VII/VIII, rostral to the otocyst (ot); crest cells from prorhombomere C are migrating into arches 3 and 4 and starting to form the primordia of cranial ganglia IX and X, caudal to the otocyst. d, diencephalon; fb, forebrain; hn, Hensen’s node; ht, heart; mb, midbrain; oc, occipital region of neural fold/tube; p, pharynx; r1, rhombomere 1 of the hindbrain; t, telencephalon; Rathke’s pouch is indicated by a small arrow on B. (A and B, From Morriss-Kay and Wilkie 2005; C and D, from Jiang et al 2002.)
Caudal to the segmented region of the neural tube, neural crest cells from rhombomere 8 migrate with the occipital myotome-derived mesenchyme to form the hypoglossal cord, eventually differentiating to form the connective tissue (neural crest) and musculature of the tongue. No sensory ganglia are formed from the occipital neural crest in human embryos. The origin of the rhombomeres and brain regions in human embryos is given in Figures 24.2 and 24.12.
Ectodermal placodes also make important contributions to the developing head. There is good evidence to suggest that all cranial placodes originate from a continuous pre-placodal territory, the ectodermal ring, around the periphery of the early neural plate, distinct from neural and neural crest cell precursors; this territory is then subdivided into separate placodes, each with its distinct developmental fate (Bailey & Streit 2006). During neurulation the placodes are segregated into the surface ectoderm, moving within the epithelium to their final positions.
The first transverse division to form in the early embryonic brain is the preotic sulcus (Figs 35.1A, 35.2A), which defines the boundary between prorhombomeres A and B. This early division is of major functional and organizational significance, since it separates the regions of origin of the skeletogenic neural crest populations that make a major contribution to the skull from those that form skeletal structures in the neck (hyoid bone and larynx). During cranial neurulation the prorhombomeres undergo subdivision so that by the time the brain region of the neural tube closes, seven rhombomeres can be clearly distinguished; the occipital region (adjacent to the four occipital somites) remains unsegmented (Fig. 35.2B). By analogy with the segmented rhombencephalon, it is also referred to as prorhombomere D/rhombomere A. The segmental organization of the embryonic head caudal to prorhombomere A is related to the expression of evolutionarily conserved HOX genes (written Hox in the mouse) that have their rostral boundaries at specific rhombomere divisions; they are also expressed in the corresponding neural crest cells (reviewed by Santagati & Rijli 2003; see also Fig. 24.16). Genetic experiments in the mouse and other vertebrates have shown that skeletal patterning of the pharyngeal arch neural crest cells depends on the absence of Hox gene expression in the cells migrating into the first arch, and the expression of Hoxa2 in the second arch crest cells. Investigation of Hox gene functions in arches 3–6 has not revealed clear skeletal patterning functions.
EMBRYONIC PHARYNX AND PHARYNGEAL ARCHES
Each pharyngeal arch consists of an epithelial covering, ectoderm and endoderm, filled with mesenchyme which is mainly of neural crest origin, with some contribution from mesoderm (primitive streak-derived mesenchyme) (Fig. 35.3A,B; Table 35.1). The neural crest cells of each arch form a skeletal element and associated connective tissue; they also give rise to the walls of an aortic arch blood vessel which is lined with endothelium derived from angiogenic mesenchyme. Paraxial mesoderm forms the muscle associated with each pharyngeal arch and motor and sensory innervation derives from arch-specific cranial nerves. For an overview of the organization of these tissues in the pharynx, see Fig. 12.4; for their fate, see Figs 35.3, 35.5–35.8. Since the organization of the embryonic pharynx and related structures is similar in all vertebrate embryos, this phase has been termed ‘pharyngula’.
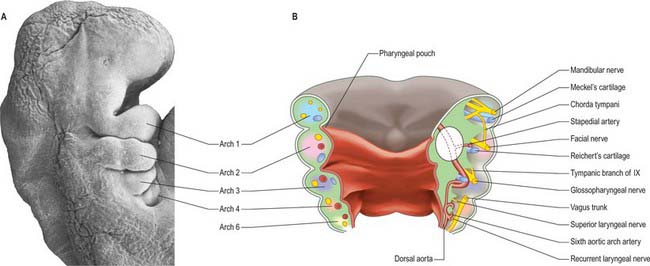

SKELETAL ELEMENTS OF THE PHARYNGEAL ARCHES
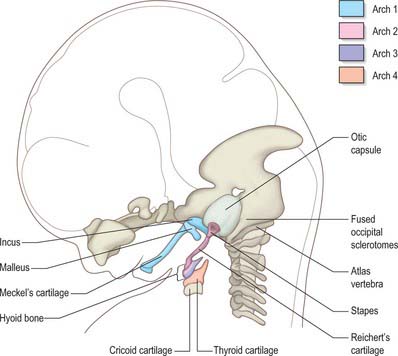
The skeletal elements of each arch are formed by condensation of neural crest-derived mesenchyme, which subsequently chondrifies in part or all of its length. Where chondrogenesis is complete (first and second arch cartilages), the element extends dorsally until it comes into contact with the cranial base lateral to the hindbrain (Fig. 35.5). An arch cartilage may remain as cartilage, undergo endochondral ossification, or become ligamentous, or a combination of these (Fig. 35.6).
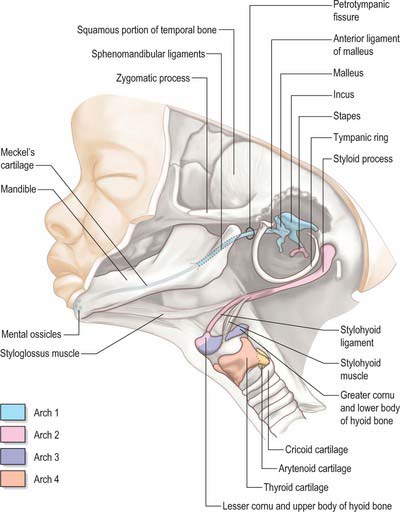
The first arch cartilage, Meckel’s cartilage, forms the embryonic jaw skeleton. After the mandible forms by intramembranous ossification of the neural crest-derived mesenchyme lateral to it, Meckel’s cartilage degenerates but its sheath persists as the anterior malleolar and sphenomandibular ligaments. Its proximal end undergoes endochondral ossification to form two of the middle ear bones, the incus and malleus. These are homologous with the quadrate and articular bones of reptiles, and the joint between them is evolutionarily derived from the reptilian jaw articulation. In reptiles, the quadrate forms from the caudal end of the palatopterygoquadrate cartilage; the mammalian evolutionary derivatives of the palatopterygoquadrate are thought to include part of the greater wing of the sphenoid bone and the roots of its pterygoid plates, in addition to the incus. The portion of Meckel’s cartilage that extends from the mental foramen almost to the site of the future mandibular symphysis probably becomes ossified and incorporated into the intramembranous mandibular bone, and the remainder of the cartilage is ultimately absorbed.
MUSCLES OF THE PHARYNGEAL ARCHES
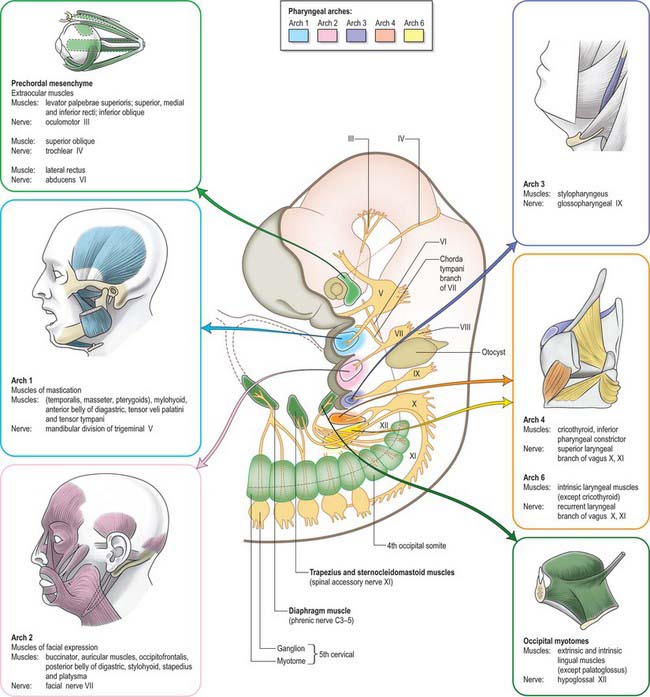
The striated muscle of each arch (sometimes termed branchiomeric) is derived from the rostral continuation of the paraxial mesoderm. The paraxial mesoderm of the head rostral to the occipital region is unsegmented, and the suggestion that a segmental pattern of seven cranial somitomeres exists is not generally accepted today. In the occipital region of the head, epithelialization takes place to form four pairs of somites; these are similar to those of the trunk. Myoblasts migrate from the paraxial mesoderm to sites of future muscle differentiation and form premuscle condensations prior to the development of any skeletal elements. The pattern of primary myotube alignment for any one muscle is specified by the surrounding neural crest-derived mesenchyme and is not related to the source of the myoblasts. The rate and pattern of muscle maturation are closely associated with the development of the skeletal elements but remain unattached until an appropriate time. Figs 35.3 and 35.7 show the muscle masses of each arch, their innervation and their derivatives in the adult.
The muscle mass of the mandibular part of the first arch forms tensor tympani, tensor veli palatini, mylohyoid, anterior belly of digastric, and the masticatory muscles (Fig. 35.7). Tensor tympani retains its connection with the skeletal element of the arch through its attachments to the malleus, and tensor veli palatini remains attached to the base of the medial pterygoid process – which may be derived from the dorsal cartilage of the first arch. However, the masticatory muscles become attached to the mandible, which is mainly a dermal bone. All of these muscles are supplied by the mandibular nerve, the mixed nerve of the first arch.
The muscles of the second arch migrate widely, but retain their original nerve supply from the facial nerve: migration is facilitated by the early obliteration of some of the first groove (cleft) and pouch. Stapedius, stylohyoid and posterior belly of digastric remain attached to the hyoid skeleton, but the facial musculature, platysma, auricular muscles and epicranius all lose connection with it (Fig. 35.7).
The muscle masses from arches 3 and 4 are adapted to form the musculature of the pharynx, larynx and soft palate. Stylopharyngeus is a third arch muscle, cricothyroid develops in the fourth arch (Fig. 35.7), and the rest of the laryngeal muscles are derived from the sixth arch. The precise origin of the remaining palatal muscles and the pharyngeal constrictors is uncertain. Palatoglossus, a muscle of the palate, is innervated by the cranial part of the accessory nerve and may derive from the lower arches.
NERVES OF THE PHARYNGEAL ARCHES
The nerves associated with each arch arise from the adjacent hindbrain (Fig. 35.7; see also Figs 12.4, 24.18). They are cranial nerves V (trigeminal), VII (facial), IX (glossopharyngeal), X (vagal) and XI (accessory). The motor nerves extend from the basal plate of the hindbrain to innervate the striated muscle of the arches: they are termed special branchial (visceral) efferent nerves because they innervate pharyngeal (branchiomeric) musculature. Sensory nerves extend peripherally and centrally from cranial ganglia that are formed partly from neural crest and partly from cells that delaminate from epipharyngeal placodes; they convey general and special somatic afferent axons. Each arch is innervated by a mixed nerve, but the nerves of arches 1–3 also have a purely sensory branch that innervates the arch rostral to its ‘own’ arch; this sensory nerve is called the pretrematic branch, because it extends rostral to the cleft or trema between the two arches (Figs 35.3, 35.7). Hence, the nerves to the mandibular arch include the mandibular division of the trigeminal nerve, which is mixed, and the chorda tympani and greater petrosal nerves, purely sensory pretrematic branches of the facial nerve. The maxillary branch of the trigeminal and the tympanic branch of the glossopharyngeal are also considered to be pretrematic nerves. The ophthalmic branch of the trigeminal nerve, which supplies the frontonasal area, is not an arch nerve; in fishes its ganglion is separate from the first arch nerve ganglion.
The vagus nerve, together with some fibres from the cranial part of the accessory nerve, supplies arches 3 (pharyngeal muscles), 4 and 6 (Fig. 35.7). The recurrent laryngeal branch initially loops under the sixth arch artery on both sides, but the asymmetric changes to the vascular system affect the symmetry of this nerve. On the right, where the sixth aortic arch disappears beyond the right pulmonary artery, the recurrent laryngeal nerve loops under the fourth arch-derived subclavian artery; on the left it loops round the ductus arteriosus (ligamentum arteriosum in the adult). Hence, the right and left branches of this nerve have asymmetric courses in the adult. Further details of cranial nerve development and composition are provided in Chapter 24.
BLOOD VESSELS OF THE PHARYNGEAL ARCHES
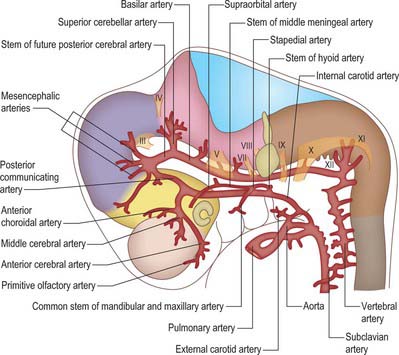
The aortic arches initially develop by vasculogenesis, soon after neural crest cells have invaded the early pharyngeal arches. Angiogenic mesenchyme forms the endothelial lining of the vessels and neural crest contributes to the outer layers of the walls (Jiang et al, 2000). The first aortic arch artery is part of the original vascular circuit that links the truncus arteriosus of the heart to the paired dorsal aortae, blood returning to the heart via the allantoic, vitelline and common cardinal veins (see Figs 13.1, 59.8). As the heart descends relative to the forebrain and other rostral structures, the aortic sac gives rise to paired aortic arches at successively more caudal levels, each of which passes laterally on each side of the pharynx to join the dorsal aortae. The dorsal aortae are not invested by neural crest cells, and nor are the more distal (cranial) parts of the carotid arteries, which form by angiogenesis. The whole complement of aortic arches never coexists – the first aortic arch degenerates through remodelling of the cranial arteries before the sixth is formed.
When the first and second aortic arch arteries begin to regress, the supply to the corresponding pharyngeal arches is derived from a transient ventral pharyngeal artery which terminates by dividing into mandibular and maxillary branches. The second aortic arch artery also degenerates, remaining only as the stem of the stapedial artery. The early vessel anastomoses with the ventral pharyngeal artery passing as it does through the mesenchyme of the stapedial cartilage, which condenses around it, forming the foramen of the stapes. The fully developed stapedial artery possesses three branches, mandibular, maxillary and supraorbital, which follow the divisions of the trigeminal nerve (Fig. 35.8). The mandibular and maxillary branches diverge from a common stem.
Further development of the third, fourth and sixth aortic arches (Fig. 35.9) produces the main arteries to the head and the great vessels arising from the heart. The external carotid artery first appears as a sprout, which extends cranially from the aortic sac close to the ventral end of the third arch artery. The common carotid arises from an elongation of the adjacent part of the aortic sac, and the third arch artery becomes the proximal part of the internal carotid artery. The fourth aortic arch on the right forms the proximal part of the right subclavian artery, whereas the corresponding vessel on the left constitutes the arch of the definitive aorta between the origins of the left common carotid and left subclavian arteries.
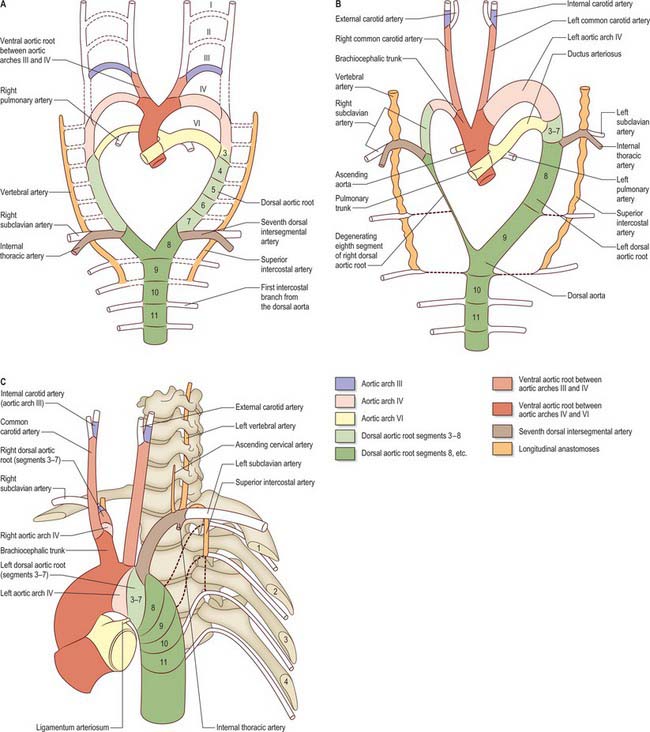
When the definitive ophthalmic artery differentiates as a branch from the terminal part of the internal carotid artery, it communicates with the supraorbital branch of the stapedial artery which distally becomes the lacrimal artery. The latter retains an anastomotic connection with the middle meningeal artery. The dorsal stem of the original second arch artery remains as one or more caroticotympanic branches of the internal carotid artery.
From its inception, the sixth aortic arch is associated with the developing lung buds. Initially, each bud is supplied by a capillary plexus from the aortic sac; as the sixth aortic arch develops, it becomes the channel for blood from the aortic sac to the developing lung buds as well as to the corresponding dorsal aorta, the latter being the main channel. Soon after formation of the sixth aortic arch, the outflow tract of the heart is divided by an influx of neural crest cells which form the spiral aorticopulmonary septum (Jiang et al 2000) which separates the aortic sac into the pulmonary trunk and ascending aorta; the pulmonary trunk attains alignment with the left sixth aortic arch (see p. 1025). The part of the sixth aortic arch between the pulmonary trunk and the dorsal aorta becomes the main vascular channel on the left, and is defined as the ductus arteriosus when the equivalent vessel on the right side is lost. Loss of the right sixth aortic arch may be related to the changes in blood flow dynamics consequent on division of the outflow tract. The caudal part of the right dorsal aorta (close to its conjunction with the left) also degenerates, separating the definitive aorta from the developing subclavian artery (Fig. 35.9B). The ductus arteriosus remains the main channel for blood to pass from the right ventricle to the descending aorta until the lungs and their associated blood vessels expand at birth. After birth, the ductus arteriosus is functionally closed by contraction of the circular muscle of the tunica media (see Ch. 59) (Fig. 35.9C).
PHARYNGEAL ECTODERM AND CLEFTS
Surface ectoderm lines the roof of the embryonic pharynx up to and including the adenohypophysial pouch (for the early development of the pituitary gland, see page 367 of Chapter 24). It also completely clothes the first arch covering the lateral walls and floor of the pharynx, unlike the more caudal arches which are dependent on the proximity of pharyngeal endoderm for their development. The external surface ectoderm of the first arch ultimately produces the keratinized stratified squamous epithelium of the epidermis including hair follicles, sweat and sebaceous glands, and the specialized epithelium of the vermilion of the lip. The first pharyngeal cleft is obliterated ventrally; its dorsal end deepens to form the external acoustic meatus and the external surface of the tympanic membrane (Fig. 35.4B,C); the ectoderm giving rise to the lining epidermis and ceruminous glands. The ectodermal covering of the oral aspect of the first arch forms the mucous membranes of the palate, oral cavity and anterior part of the tongue and the ectodermal components of the teeth, salivary and mucous glands. Three auricular hillocks form on arches 1 and 2, each side of the first pharyngeal groove beginning at stage 15; they form the auricle of the external ear (see Ch. 38).
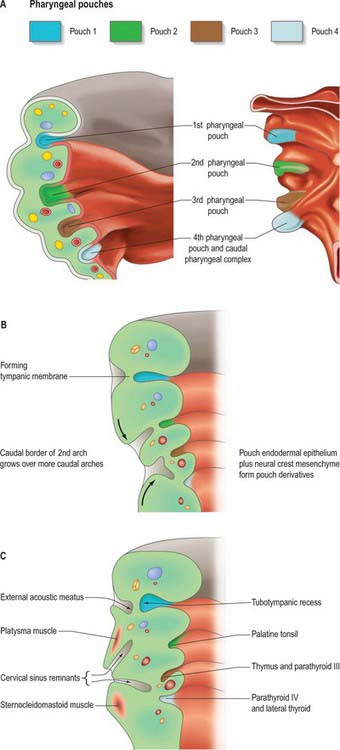
Fig. 35.4 Pharyngeal pouch development. A. The arrangement of the early pharyngeal pouches: on the left, the internal aspect of the pharyngeal floor viewed from above; on the right, the external aspect of the pharyngeal floor viewed from below. B. Coronal section of the left side of the pharynx at stage 18, showing changes to the pharyngeal pouches internally and pharyngeal clefts externally. C. Coronal section of the left side of the pharynx at stage 19. See also Fig 35.18 for further development.
The external contours of the arches and clefts are modified as the skeletal and muscular elements develop. During the fifth week, pharyngeal clefts 2 and 3+4 form the rostral and caudal parts of a retrohyoid depression, the cervical sinus (Fig. 35.4C). Cranially, the sinus is bounded by the hyoid arch; dorsally, by a ridge produced by ventral extensions from the occipital myotomes and by mesenchyme that develops into the sternocleidomastoid and trapezius muscles. Caudally, the smaller epipericardial ridge separates the sinus from the pericardium. Fusion of the hyoid arch with the cardiac elevation covers the cervical sinus, excluding the arches 3–6 from contributing to the skin of the neck; it also results in platysma muscle, bounded both superficially and deep with superficial fascia, passing along the neck to the anterior thoracic wall.
Thickened patches of ectoderm, the epibranchial placodes, appear at the dorsal ends of the pharyngeal grooves. They are closely related to the developing ganglia of the facial, glossopharyngeal and vagus nerves, to which they contribute. Together with dorsolateral and suprabranchial placodal cells, the epibranchial placodes also contribute to the trigeminal and vestibulocochlear ganglia (see Figs 12.4, 24.18).
PHARYNGEAL ENDODERM AND POUCHES
The first four pharyngeal pouches appear in sequence craniocaudally during stages 10–13. The rostral pharynx is broad and dorsoventrally compressed (see Fig. 35.18). Laterally, the endoderm of the pouches approaches the ectoderm of the pharyngeal clefts to form thin closing membranes (Fig. 35.4). The approximating ectoderm and endoderm between the first cleft and pouch form the outer and inner surfaces of the tympanic membrane. The ventral end of the first pouch is obliterated, but its dorsal end persists and expands as the head enlarges. This, together with the adjoining lateral part of the pharynx and possibly with a contribution from the dorsal part of the second pharyngeal pouch, constitutes the tubotympanic recess. The recess forms the middle ear cavity, the pharyngotympanic tube and their extensions. Expansion of the middle ear cavity around the ear ossicles occurs late during fetal life, by breakdown of the mesenchyme around the ossicles so that the cavity comes to surround them except at their attachments to the tympanic membrane and fenestra ovalis (fenestra vestibuli, oval window).
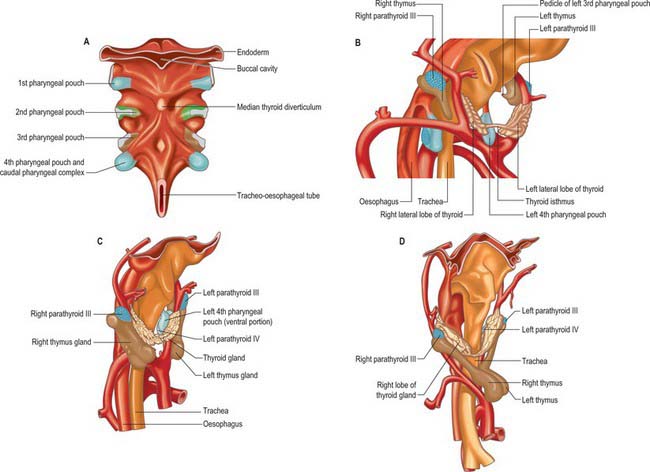
The ectoderm of the pharyngeal clefts and the endoderm of the pouches become increasingly separated by mesenchyme. The blind recesses of the second, third and fourth pouches are prolonged dorsally and ventrally as angular, wing-like diverticula (see Fig. 35.18); the pouch endoderm thickens and evaginates into localized neural crest-derived mesenchymal condensations. The ventral part of the second pouch is the focus of lymphoid development as the palatine tonsil. A generalized ring of lymphoid tissue, the lymphatic ring, develops in this region. The third pouch gives rise to the thymus ventrally and the parathyroid III dorsally, whereas the fourth pouch produces the parathyroid IV and an ultimobranchial body. The dorsal and ventral portions of the fourth pouch, together with the lower ultimobranchial body, are collectively termed the caudal pharyngeal complex.



