Chapter 41 Plant description, morphology and anatomy
Plant form ranges from unicellular plants—for example, yeasts and some green algae—to the strongly differentiated higher plants. Examples of pharmaceutical interest may be found in most of the larger groups and a quick perusal of the families involved in Chapter 5 of this textbook illustrates this point.
As indicated in Chapter 2, for convenience of study, drugs may be arranged not only according to families and chemical constituents, but also into such morphological groups as barks, roots, leaves, seeds, etc. Some drugs constitute more than one morphological part—for example, whole herbs and commercial ‘roots’, which may consist of both rhizomes and roots.
LEAVES AND TOPS (′HERBS′)
Structure of the aerial stem
The primary stem (Fig. 41.1A) shows the following structure: epidermis, cortex, medullary rays, medulla and a vascular system taking the form of a dictyostele. The epidermis is composed of a single layer of compactly arranged cells and bears stomata. The cortex is usually parenchymatous, the outer layers of cells in aerial stems containing chloroplasts. The layers of cortex cells immediately underlying the epidermis may be collenchymatous, constituting a hypodermis. The endodermis is usually not well-differentiated in aerial stems, although a layer of cells containing starch (starch sheath) and corresponding in position to the endodermis may be defined. Underground stems often resemble roots in showing a more or less well-differentiated endodermis with characteristic Casparian strips (thickenings).
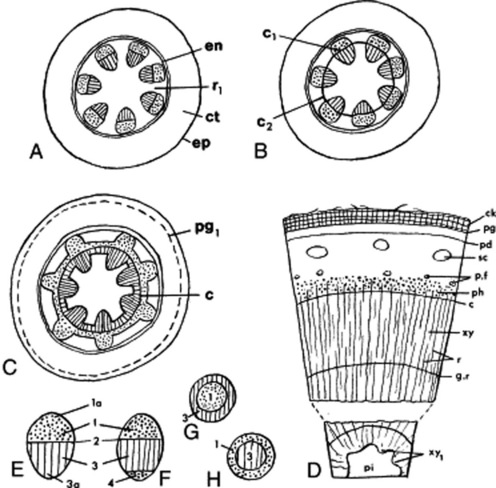
The vascular bundles of the dictyostele are usually collateral, but are in some cases bicollateral (Cucurbitaceae, Solanaceae, Convolvulaceae) (Fig. 41.1E–H). The xylem is differentiated centrifugally and the protoxylem is endarch; the phloem is differentiated centripetally and the protophloem is exarch (cf. the root). The differentiation, in dicotyledons, is usually incomplete, so that a zone of meristematic cells (the intrafascicular cambium) separates the primary vascular tissues. Such a bundle is described as open, in contrast to the closed bundle typical of monocotyledons. In the bicollateral bundle the intrafascicular cambium occurs between the xylem and the outer phloem group.
Secondary thickening is initiated by tangential divisions in the intrafascicular cambium. The daughter cells cut off on the inner side differentiate into xylem and those cut off to the outside into phloem. The amount of secondary xylem produced in both stems and roots, in general, exceeds the amount of secondary phloem. As the process of secondary thickening of the stem proceeds, its dictyostele is converted into a solid cylinder of secondary tissues. The intrafascicular cambia become linked to form a continuous cambial cylinder by the development of interfascicular cambia in the ray tissue (Fig. 41.1B, C). The cambial activity may spread out from the intrafascicular cambia across the rays, or in other cases cambial activity may originate at a median point in the ray and then by lateral extension from both intrafascicular and interfascicular cambia the cambial cylinder may be completed.
In woody perennials the cambial divisions are arrested during the winter but are renewed each spring. The xylem produced at different seasons varies in texture. The spring wood is characterized by abundance of relatively thin-walled large conducting elements; the autumn wood, by a high proportion of thick-walled mechanical elements such as wood fibres. A similar alteration between sieve tissue and phloem fibres may occur in the secondary phloem. With increase in girth the central core of xylem may become non-functional, dark in colour and packed with metabolic byproducts forming a heartwood or duramen. Sandalwood is the heartwood of Santalum album and is packed with volatile oil. The blocking of the vessels in the formation of heartwood occurs by the development of tyloses (see Fig. 42.6P).
The secondary increase in diameter of the vascular cylinder is accompanied by changes in the outer tissues. The epidermis and part or all of the primary cortex may be shed. A phellogen may arise in the epidermis, cortex or pericycle and give rise externally to cork and internally to a variable amount of phelloderm (Fig. 41.1D).
For the investigation of the anatomy of stems, transverse sections and radial and tangential longitudinal sections should be prepared from the drug previously moistened or soaked. For a study of the individual elements, disintegrated material should be used (see Chapter 43).
BARKS
A young bark (Fig. 41.1) is composed of the following tissues.
The cork cambium or phellogen may arise in the epidermis (e.g. willow), primary cortex or pericycle. The phellogen produces on its outer side cork, and on its inner side chlorophyll containing suberized cells which form the secondary cortex or phelloderm. These three layers are known as the periderm. If the cork cambium develops in or near the pericycle, a part of the whole of the primary cortex will lie outside the cork and will be gradually thrown off. Lenticels replace stomata for purposes of gaseous exchange; and as the cork increases, the amount of chlorophyll-containing tissue decreases.
The natural curvature of the bark increases when the bark is removed from the tree and dried. Large pieces of trunk bark, especially if subjected to pressure, may be nearly flat. Terms used to describe the curvature are illustrated in Fig. 41.2. Some commercial barks (e.g. cinnamon and quillaia) consist of the inner bark only. In quillaia the dark patches often found on the outer surface are known as rhytidome (literally, ‘a wrinkle’). This term is applied to plates of tissue formed in the inner bark by successive development of cork cambia.
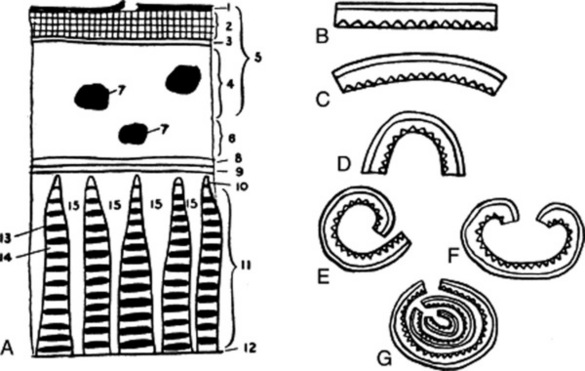
Barks may be described under the following headings.
WOODS
Although few drugs consist solely of wood, no description of a stem or root is complete without an account of its wood. Wood consists of the secondary tissues produced by the cambium on its inner surface. The cells composing these tissues—the vessels, tracheids, wood fibresand parenchyma—are not necessarily all lignified. In some cases (e.g. the wood of belladonna root) non-lignified elements predominate. The distribution of the lignified elements may be ascertained by treating smoothed transverse, radial and tangential surfaces or sections with phloroglucinol and hydrochloric acid. In trees, the cells of the old wood frequently become coloured as they fill with waste products such as resins, tannins and colouring matters. This central region is called the heartwood, while the outer wood, which still retains its normal appearance and functions, is called the sapwood. Commercial guaiacum wood and logwood consist of heartwood.
Woods may be described under the following headings.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


