INTRODUCTION
Cultivation is the process of propagating organisms by providing the proper environmental conditions. Growing microorganisms are making replicas of themselves, and they require the elements present in their chemical composition. Nutrients must provide these elements in metabolically accessible form. In addition, organisms require metabolic energy to synthesize macromolecules and maintain essential chemical gradients across their membranes. Factors that must be controlled during growth include the nutrients, pH, temperature, aeration, salt concentration, and ionic strength of the medium.
REQUIREMENTS FOR GROWTH
Most of the dry weight of microorganisms is organic matter containing the elements carbon, hydrogen, nitrogen, oxygen, phosphorus, and sulfur. In addition, inorganic ions such as potassium, sodium, iron, magnesium, calcium, and chloride are required to facilitate enzymatic catalysis and to maintain chemical gradients across the cell membrane.
For the most part, the organic matter is in macromolecules formed by the introduction of anhydride bonds between building blocks. Synthesis of the anhydride bonds requires chemical energy, which is provided by the two phosphodiester bonds in adenosine triphosphate (ATP; see Chapter 6). Additional energy required to maintain a relatively constant cytoplasmic composition during growth in a range of extracellular chemical environments is derived from the proton motive force. The proton motive force is the potential energy that can be derived by passage of a proton across a membrane. In eukaryotes, the membrane may be part of the mitochondrion or the chloroplast. In prokaryotes, the membrane is the cytoplasmic membrane of the cell.
The proton motive force is an electrochemical gradient with two components, a difference in pH (hydrogen ion concentration) and a difference in ionic charge. The charge on the outside of the bacterial membrane is more positive than the charge on the inside, and the difference in charge contributes to the free energy released when a proton enters the cytoplasm from outside the membrane. Metabolic processes that generate the proton motive force are discussed in Chapter 6. The free energy may be used to move the cell, to maintain ionic or molecular gradients across the membrane, to synthesize anhydride bonds in ATP, or for a combination of these purposes. Alternatively, cells given a source of ATP may use its anhydride bond energy to create a proton motive force that in turn may be used to move the cell and to maintain chemical gradients.
To grow, an organism requires all of the elements in its organic matter and the full complement of ions required for energetics and catalysis. In addition, there must be a source of energy to establish the proton motive force and to allow macromolecular synthesis. Microorganisms vary widely in their nutritional demands and their sources of metabolic energy.
SOURCES OF METABOLIC ENERGY
The three major mechanisms for generating metabolic energy are fermentation, respiration, and photosynthesis. At least one of these mechanisms must be used if an organism is to grow.
The formation of ATP in fermentation is not coupled to the transfer of electrons. Fermentation is characterized by substrate phosphorylation, an enzymatic process in which a pyrophosphate bond is donated directly to adenosine diphosphate (ADP) by a phosphorylated metabolic intermediate. The phosphorylated intermediates are formed by metabolic rearrangement of a fermentable substrate such as glucose, lactose, or arginine. Because fermentations are not accompanied by a change in the overall oxidation-reduction state of the fermentable substrate, the elemental composition of the products of fermentation must be identical to those of the substrates. For example, fermentation of a molecule of glucose (C6H12O6) by the Embden-Meyerhof pathway (see Chapter 6) yields a net gain of two pyrophosphate bonds in ATP and produces two molecules of lactic acid (C3H6O3).
Respiration is analogous to the coupling of an energy-dependent process to the discharge of a battery. Chemical reduction of an oxidant (electron acceptor) through a specific series of electron carriers in the membrane establishes the proton motive force across the bacterial membrane. The reductant (electron donor) may be organic or inorganic (eg, lactic acid serves as a reductant for some organisms, and hydrogen gas is a reductant for other organisms). Gaseous oxygen (O2) often is used as an oxidant, but alternative oxidants that are used by some organisms include carbon dioxide (CO2), sulfate (SO42−), and nitrate (NO3−).
Photosynthesis is similar to respiration in that the reduction of an oxidant via a specific series of electron carriers establishes the proton motive force. The difference in the two processes is that in photosynthesis, the reductant and oxidant are created photochemically by light energy absorbed by pigments in the membrane; thus, photosynthesis can continue only as long as there is a source of light energy. Plants and some bacteria are able to invest a substantial amount of light energy in making water a reductant for carbon dioxide. Oxygen is evolved in this process, and organic matter is produced. Respiration, the energetically favorable oxidation of organic matter by an electron acceptor such as oxygen, can provide photosynthetic organisms with energy in the absence of light.
NUTRITION
Nutrients in growth media must contain all the elements necessary for the biologic synthesis of new organisms. In the following discussion, nutrients are classified according to the elements they supply.
As already mentioned, plants and some bacteria are able to use photosynthetic energy to reduce carbon dioxide at the expense of water. These organisms are referred to as autotrophs, creatures that do not require organic nutrients for growth. Other autotrophic microorganisms are the chemolithotrophs, organisms that use an inorganic substrate such as hydrogen or thiosulfate as a reductant and carbon dioxide as a carbon source.
Heterotrophs require organic carbon for growth, and the organic carbon must be in a form that can be assimilated. Naphthalene, for example, can provide all of the carbon and energy required for respiratory heterotrophic growth, but very few organisms possess the metabolic pathway necessary for naphthalene assimilation. Glucose, on the other hand, can support the fermentative or respiratory growth of many organisms. It is important that growth substrates be supplied at levels appropriate for the microbial strain that is being grown: Levels that will support the growth of one organism may inhibit the growth of another organism.
Carbon dioxide is required for a number of biosynthetic reactions. Many respiratory organisms produce more than enough carbon dioxide to meet this requirement, but others require a source of carbon dioxide in their growth medium.
Nitrogen is a major component of proteins, nucleic acids, and other compounds, accounting for approximately 5% of the dry weight of a typical bacterial cell. Inorganic dinitrogen (N2) is very prevalent, comprising 80% of the earth’s atmosphere. It is also a very stable compound, primarily because of the high activation energy required to break the nitrogen–nitrogen triple bond. However, nitrogen may be supplied in a number of different forms, and microorganisms vary in their abilities to assimilate nitrogen (Table 5-1). The end product of all pathways for nitrogen assimilation is the most reduced form of the element, ammonia (NH3). When NH3 is available, it diffuses into most bacteria through transmembrane channels as dissolved gaseous NH3 rather than ionic ammonium ion (NH4+).
The ability to assimilate N2 reductively via NH3, which is called nitrogen fixation, is a property unique to prokaryotes, and relatively few bacteria are capable of breaking the nitrogen–nitrogen triple bond. This process (see Chapter 6) requires a large amount of metabolic energy and is readily inactivated by oxygen. The capacity for nitrogen fixation is found in widely divergent bacteria that have evolved quite different biochemical strategies to protect their nitrogen-fixing enzymes from oxygen.
Most microorganisms can use NH3 as a sole nitrogen source, and many organisms possess the ability to produce NH3 from amines (R—NH2) or from amino acids (RCHNH2COOH), generally intracellularly. Production of NH3 from the deamination of amino acids is called ammonification. Ammonia is introduced into organic matter by biochemical pathways involving glutamate and glutamine. These pathways are discussed in Chapter 6.
Many microorganisms possess the ability to assimilate nitrate (NO3−) and nitrite (NO2−) reductively by conversion of these ions into NH3. These processes are termed assimilatory nitrate reduction and assimilatory nitrite reduction, respectively. These pathways for assimilation differ from pathways used for dissimilation of nitrate and nitrite. The dissimilatory pathways are used by organisms that use these ions as terminal electron acceptors in respiration. Some autotrophic bacteria (eg, Nitrosomonas, Nitrobacter spp.) are able to convert NH3 to gaseous N2 under anaerobic conditions; this process is known as denitrification. Our understanding of the nitrogen cycle continues to evolve. In the mid-1990s, the anammox reaction was discovered. The reaction
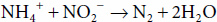
in which ammonia is oxidized by nitrite is a microbial process that occurs in anoxic waters of the ocean and is a major pathway by which nitrogen is returned to the atmosphere.
Similar to nitrogen, sulfur is a component of many organic cell substances. It forms part of the structure of several coenzymes and is found in the cysteinyl and methionyl side chains of proteins. Sulfur in its elemental form cannot be used by plants or animals. However, some autotrophic bacteria can oxidize it to sulfate (SO42−). Most microorganisms can use sulfate as a sulfur source, reducing the sulfate to the level of hydrogen sulfide (H2S). Some microorganisms can assimilate H2S directly from the growth medium, but this compound can be toxic to many organisms.
Phosphate (PO43−) is required as a component of ATP; nucleic acids, and such coenzymes as NAD, NADP, and flavins. In addition, many metabolites, lipids (phospholipids, lipid A), cell wall components (teichoic acid), some capsular polysaccharides, and some proteins are phosphorylated. Phosphate is always assimilated as free inorganic phosphate (Pi).
Numerous minerals are required for enzyme function. Magnesium ion (Mg2+) and ferrous ion (Fe2+) are also found in porphyrin derivatives: magnesium in the chlorophyll molecule, and iron as part of the coenzymes of the cytochromes and peroxidases. Mg2+ and K+ are both essential for the function and integrity of ribosomes. Ca2+ is required as a constituent of gram-positive cell walls, although it is dispensable for gram-negative bacteria. Many marine organisms require Na+ for growth. In formulating a medium for the cultivation of most microorganisms, it is necessary to provide sources of potassium, magnesium, calcium, and iron, usually as their ions (K+, Mg2+, Ca2+, and Fe2+). Many other minerals (eg, Mn2+, Mo2+, Co2+, Cu2+, and Zn2+) are required; these frequently can be provided in tap water or as contaminants of other medium ingredients.
The uptake of iron, which forms insoluble hydroxides at neutral pH, is facilitated in many bacteria and fungi by their production of siderophores—compounds that chelate iron and promote its transport as a soluble complex. These include hydroxamates (-CONH2
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


