Chapter 5 Crystalline Lens
The crystalline lens is an avascular, transparent elliptic structure that aids in focusing light rays on the retina. The lens is located within the posterior chamber, anterior to the vitreous chamber and posterior to the iris (Figure 5-1). The lens is suspended from the surrounding ciliary body by zonular fibers. It is malleable, and ciliary muscle contraction can cause a change in lens shape, increasing the dioptric power of the eye. The mechanism that causes an increase in lens power is accommodation, which allows near objects to be focused on the retina.
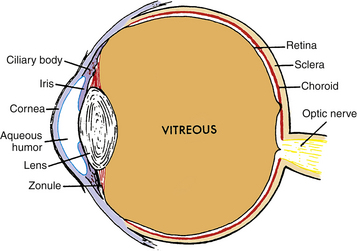
FIGURE 5-1 Diagram showing relationship of the lens and zonule to other ocular structures.
(From Paterson CA, Delamere NA: The lens. In Hart WM Jr, editor: Adler’s physiology of the eye, ed 9, St Louis, 1992, Mosby.)
The posterior lens surface is attached to the anterior vitreous face by the hyaloid capsular ligament, a circular ring adhesion. Within this ring is a potential space, the retrolental space (of Berger), an area of nonadhesion between the vitreous and the lens (see Figure 6-12).
Lens Dimensions
The lens is biconvex, with the posterior surface having the steeper curve. The anterior radius of curvature measures 8 to 14 μm, and the posterior surface radius of curvature measures 5 to 8 μm. The centers of the anterior and posterior surfaces are called the poles, and the lens thickness is the distance from the anterior to posterior pole. The thickness of the unaccommodated lens is 3.5 to 5 mm and it increases 0.02 mm each year throughout life.1–3 The lens diameter is the nasal-to-temporal measurement and in the infant is 6.5 mm. The diameter reaches 9 mm during the teenage years and does not change significantly,4–6 although some report a small age-related increase in diameter.7 The equator is the largest circumference of the lens at a location between the two poles.
The refractive power of the unaccommodated lens is approximately 20 diopters (D) and depends on the (1) surface curvatures, (2) refractive index, (3) change in index between the lens and surrounding environment, and (4) length of the optical path. The lens has a gradient refractive index because of changes in optical density throughout the lens; the index increases from the anterior to the center of the lens and decreases toward the posterior surface. The refractive index is a factor of protein concentration within the lens fibers.8 The variations in index provide additional refractive power.9,10 The power of the lens increases in accommodation, with the maximum accommodative amplitude, 14 D, reached between ages 8 and 12 years.11 Accommodative power decreases with age, approaching zero after 50 years.12
Embryologic Development
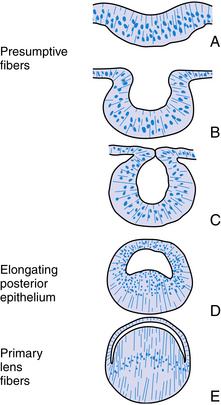
The structure of the adult lens is determined during embryologic development. The lens vesicle, the first lenslike structure observable in the developing embryo, is composed of a layer of epithelial cells that form a hollow sphere. The cells are positioned so that the apical surface lines the lumen of this sphere. The posterior cells differentiate and elongate, forming the primary lens fibers. As these fibers grow and reach the anterior cells (Figure 5-2), the center of the sphere fills. Thus the adult lens has no posterior epithelium because it was used to form these first lens fibers. During the rest of the life of the lens, cell division occurs in the germinative zone of the epithelium just anterior to the lens equator, and the cells thus formed elongate to form secondary lens fibers that are laid down outer to all earlier fibers. With age, the lens continues to grow as it forms new fibers (see Chapter 7).
Histology
Lens Capsule
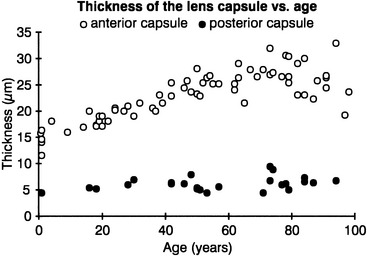
The lens capsule is a transparent envelope that surrounds the entire lens. The capsule is a basement membrane and with time becomes the thickest in the body.13 Its thickness varies with location. At the posterior pole, it is thinnest (approximately 3.5 μm) and does not appreciably increase with age; the thickness at the equator increases slightly with age, and on average is 7 μm. The capsule thickness at the anterior pole increases with age from approximately 11 to 15 μm. The annular region surrounding the anterior pole appears to be the thickest. It too increases with age from approximately 13.5 to 16 μm.14 The capsule consists primarily of collagen; it contains no elastic fibers but is highly elastic because of the lamellar arrangement of the fibers.4,15 It encloses all lens components and helps to mold the shape of the lens. The capsule would prefer to take a more spherical shape, but this tendency is counteracted by the pull from the zonular fibers. The zonular fibers insert into the capsule, merging with it in an area from the equator to near both poles. This coincides with the annular area mentioned previously. This outer superficial zone of the capsule is called the zonular lamella and consists of zonules interconnected with matrix.4 The lens capsule provides some barrier function preventing large molecules, such as albumin and hemoglobin,4 from entering the lens. The anterior lens capsule is produced by the anterior epithelium and thickens with age. The posterior lens capsule may receive some contribution from the basal membrane of lens fibers, but the thickness of the posterior capsule changes minimally throughout life (Figure 5-3).16–18
Lens Epithelium
Adjacent to the anterior lens capsule is a layer of cuboidal epithelium—the anterior lens epithelium (Figure 5-4). These cells secrete the anterior capsule throughout life and are the site of metabolic transport mechanisms. As noted, no posterior epithelium is present because it was used during embryologic development to form the primary lens fibers. The basal aspect of the epithelial cell is adjacent to the capsule, and the apical portion is oriented inward toward the center of the lens. The lateral membranes of the epithelial cells are joined by desmosomes and gap junctions.17,19–22 Although zonula occludens were once thought to join the epithelial cells,23 more recent studies report few, if any, tight junctions.18

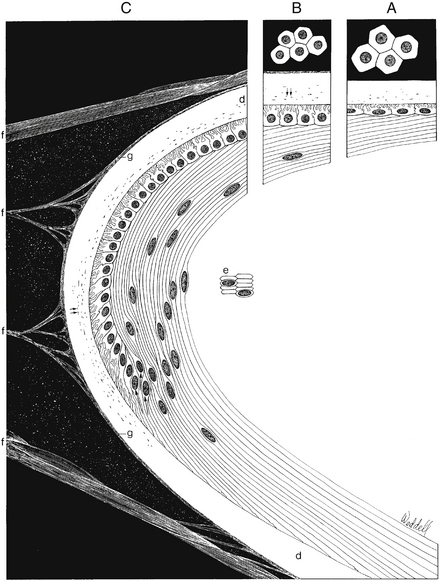
The band of cells in the preequatorial region that lies just anterior to the equator is called the germinal zone, the location of cell mitosis. Cell division continues throughout life; as each cell divides, a daughter cell migrates posteriorly towards the equator, withdraws from the cell cycle, and differentiates into a lens fiber. Each newly formed cell elongates; the basal aspect stretches toward the posterior pole and the apical aspect toward the anterior pole (Figure 5-5). This process occurs all around the equator, with fibers stretching toward the poles from all aspects of the lens periphery. As the cells in each layer elongate, the cellular nuclei move with the cytoplasm. A line drawn to connect the dots of these nuclei would have an arcuate shape toward the anterior aspect, a configuration called the lens bow (see Figure 5-2). Eventually, as it loses all cellular organelles, the elongated cell becomes a lens fiber. The anterior end of the lens fiber (the apical surface) insinuates itself between the epithelial layer and the underlying lens fibers. The new fibers are laid down outer to the older fibers; the more superficial fibers are longer than deeper fibers, and the youngest cells lie directly below the epithelium and the capsule. All fibers formed from mitosis in the germinative zone are called secondary lens fibers. Once it loses its nucleus, the mature lens fiber has lost its attachment to the basement membrane.17
Lens Fibers
Lens fiber production continues throughout life, with the new lens fibers being laid down outer to the older fibers; growth results in concentric layers of secondary lens fibers. The structure of the lens is similar to an onion; each layer of fibers approximates a layer of an onion, but then each layer is made up of adjacent fibers. A section through the equator of the lens shows that the fibers cut in cross section are mostly hexagonal in shape and arranged in concentric rings (Figure 5-6). The cross section dimensions of a fiber are approximately 3 by 9 μm.24 Each fiber has a long crescent shape, with the broad sides parallel to the lens surface and the narrow sides at an angle to the surface and located near the lens poles.

Lens fiber cytoplasm contains a high concentration of proteins, known as crystallins, which account for approximately 40% of the net weight of the fiber. The distribution and concentration of crystallins contribute to the gradient refractive index.25 The crystallin concentration varies from approximately 15% in the cortex to 70% in the nucleus.26 A cytoskeletal network of microtubules and filaments provides structure and also provides stability by being anchored to the plasma membrane.18,25 The lateral membranes have numerous and elaborate interdigitations along the fiber length that take various shapes, such as ball-and-socket and tongue-in-groove junctions, and allow for sliding between fibers4,17,18 (Figure 5-7). The fibers also are joined by desmosomes.
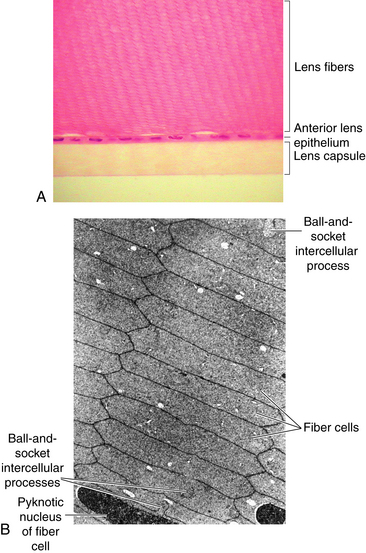
Because the lens has no vascular supply and the fibers lose their cellular organelles as they age, some cell-to-fiber and fiber-to-fiber mechanism of communication is necessary. There is an extensive network of gap junctions throughout the lens along the lateral fiber membranes to account for the facility with which nutrients and ions move within the lens.17,27 These gap junctions have a different packing arrangement and different protein connexins, forming the channel than do the typical gap junctions.21 The gap junctions are not evenly distributed throughout the lens, with few near the poles, more toward the equator, and seemingly fewer junctions in deeper layers.13,21,22 Micropinocytic vesicles at apical and basal aspects of fiber membranes and significant areas of membrane fusion also allow movement of material from fiber to fiber, contributing to communication between fibers in addition to the gap type of junctions described.13,20,22,28
Epithelium-Fiber Interface
The border between the apical membrane of the anterior epithelium and the apical membrane of the elongating fiber is known as the epithelium-fiber interface (EFI). Nutrients and ions are exchanged across the EFI. It was once assumed that such movement was facilitated by gap junctions, but disagreement now exists on whether gap junctions are present.19,22,27,28 Gap junctions are usually found on the lateral cell membrane, and the EFI involves apical surfaces. Few true gap junctions have been visualized in tissue preparations. Minimal coupling occurs between the epithelium and fibers in the central zone (i.e., near the poles and sutures), but such junctions increase toward the germinative zone.21 Pinocytosis does occur at this interface, facilitating exchange.18
Divisions of Lens
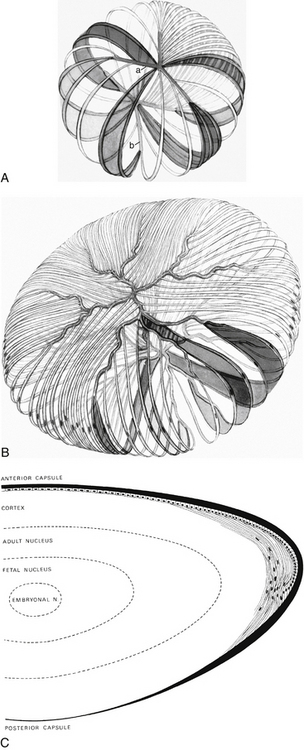
The primary lens fibers from the elongating posterior epithelium form the very center of the lens, the embryonic nucleus, and all subsequent lens fibers are laid down outer to this core. Cell mitosis then begins in the preequatorial region of the epithelium, the new cell migrates toward the equator, and then elongates, forming a lens fiber. All such fibers formed are secondary lens fibers. The fetal nucleus includes the embryonic nucleus and the fibers surrounding it that are formed before birth. The adult nucleus is considered to include the embryonic and fetal nuclei and the fibers formed from birth to sexual maturation. The lens cortex contains the fibers formed after sexual maturation (Figure 5-8, C). The cortex can be divided into superficial, internal, and deep zones,13 but these are arbitrary divisions and generally are not clinically significant. Some consider the fibers formed before sexual maturation the “juvenile nucleus,” those added before middle age the “adult nucleus,” and the remaining fibers the “cortex.”25 The lens cortex has the lowest and the embryonic nucleus has the highest index of refraction.
Lens Sutures
As the lens fibers reach the poles they meet with the other fibers in their layer, forming a junction known as a suture. The anterior suture is formed by the joining of the apical aspects of the fibers, and the posterior suture is formed by the joining of the basal aspects.4,13 The secondary fibers formed during embryologic development meet in three branches, forming Y sutures. The anterior suture is an upright-Y shape and the posterior suture an inverted-Y shape (see Figure 5-8, A).18 As growth continues and the lens becomes larger, the sutures become asymmetric and dissimilar. The limbs of the anterior and posterior sutures are offset, and the complexity of the sutures contributes to lens transparency.29 The sutures formed after birth are more stellate shaped; sutures formed through early adulthood have 6 to 9 branches; and 9 to 15 complex branched stars are formed in middle to old age29 (see Figure 5-8, B).
Zonules (of Zinn)
The lens is attached to the ciliary body by a group of threadlike fibers, the zonules (of Zinn), or the suspensory ligament of the lens (Figure 5-9). The fibers belong to a category termed microfibrils. The macromolecular composition that accounts for the remarkable extensibility of the zonules is complex and still undetermined.15,30 The zonules appear to be formed of extracellular matrix, that includes fibrillin and elastin, both of which have a role in the synthesis of elastic fibers.31 However, biomolecular analysis indicates that there are no true elastic fibers present in the zonules.31
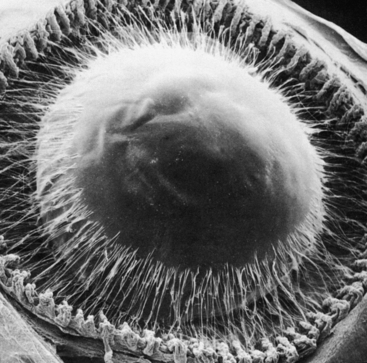
The fibers arise from the basement membrane of the nonpigmented ciliary epithelium in the pars plana and from the valleys between the ciliary processes in the pars plicata.4 They form two columnlike structures on both sides of a ciliary process and end at the lens capsule (see Figure 3-12). Most fibers attach to the lens capsule at the preequatorial and postequatorial regions; few attach directly at the equator.32 The zonules are interwoven into the components of the capsule. Those that attach to the lens are known as primary zonules. Secondary zonules join the primary zonules with each other or connect processes to one another or to the pars plana, and “tension” fibers anchor the primary zonules to the ciliary valleys to form a fulcrum.32
Accommodation
When the emmetropic eye is viewing a distant object, the ciliary muscle is relaxed, the diameter of the ciliary ring is relatively large, and the zonules are in a stretched configuration exerting tension on the lens capsule holding the lens in the unaccommodated state, such that the image lies on the retina. When a near object is to be focused on the retina, an increase in the refractive power of the eye must occur. This increase in power is called accommodation and is accomplished by a change in lens shape brought about by contraction of the ciliary muscle. Von Helmholtz33 is credited with determining that the following occur during accommodation:
These factors result in a thickened lens with more sharply curved surfaces and thus increased lens power. The stimulus that initiates the accommodative mechanism is retinal blur.34 The accommodative mechanism is dependent on cone stimulation with little influence by rods.35
The lens becomes more sharply curved, and because of the thickness variation of the capsule, a bulge is most apparent at the center of the anterior surface.36,37 The posterior surface increases in curvature only slightly; however, the steepness of the anterior surface does not become greater than that of the posterior surface.12 As the lens thickens axially, the equatorial diameter decreases, and the anterior lens pole moves toward the cornea. It was previously thought that there was no posterior lens movement, but a small amount of movement of the posterior pole in a posterior direction has been identified.8,38,39 The curvature of the internal surfaces, seen at the zones of discontinuity and the boundaries of the nuclei, mimics the changes in the surface curvatures and contributes to the increase in the total dioptric power.8 This thickening of the lens from anterior to posterior occurs in the nuclear region, but the thickness of the lens cortex remains unchanged.40 The changes occurring in accommodation described in 1855 by Helmholtz have been demonstrated using current technology.5,41
The vitreous has a passive role in accommodation, probably serving only as support for the lens.42 During ciliary muscle contraction, the choroid is pulled forward slightly, perhaps aiding in the correct orientation of the photoreceptors in relation to the entrance pupil. Contraction of the ciliary body in accommodation, by decreasing the circumference of the sclera, may lead to an elongation of the axial length of the eye.39
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


