chapter 13 Cellular communication
 Cells communicate with one another either by direct cell–cell interactions or from a distance using signalling molecules.
Cells communicate with one another either by direct cell–cell interactions or from a distance using signalling molecules.
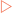
Intercellular communication
Interactions
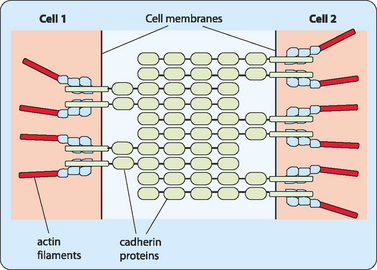
Adherens junctions are more stable interactions between cells, usually involving a specific type of CAM, the cadherins. The cytoskeletons of cells that are destined to adjoin one another in a specific tissue or organ are often joined by cadherins. In these junctions, cadherin proteins are bonded to the actin filaments that make up the cytoskeleton. The extracellular domains of cadherin proteins then interact with cadherin proteins from an adjoining cell, effectively tethering the neighbouring cells together (Fig 13-1).
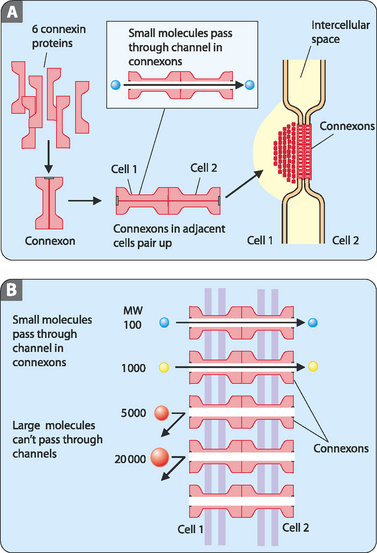
The activities of cells within an organ or tissue type often need to be tightly regulated. As discussed below, this is often controlled through receptor-mediated extracellular signalling. Adjoining cells, however, are often linked by open channels that allow for rapid cytoplasmic exchange. These gap junctions are transmembrane channels that allow for the diffusion of small molecules between the neighbouring cells (Fig 13-2A). In general, ions—small organic molecules, low-molecular-weight precursors of proteins and nucleic acids—and small signalling molecules can pass through gap junctions whereas proteins and nucleic acids cannot (Fig 13-2B). Cells that are connected by gap junctions are said to be metabolically coupled; in other words, they are nearly identical in terms of electrical and metabolic activity. Gap junctions are made up of connexins, a family of transmembrane proteins, which form a cylinder with an open channel in the centre. This assembly of connexins, the connexon, lines up with a connexon from an adjoining cell to form a continuous channel from one cell to the next.
Transport of signalling molecules
Intercellular communication mediated by signalling molecules can be broken down into three general categories—endocrine, paracrine or autocrine signalling. In endocrine signalling, a signalling molecule is released by endocrine organs or glands and moves through the general circulation until it reaches its target cell. This allows for communication from a distance. Most hormones act via endocrine signalling. Paracrine signalling involves a signalling molecule from one cell acting on neighbouring cells, allowing for short distance communication. Autocrine signalling is not, strictly speaking, intercellular communication as it involves a cell responding to signalling molecules that it produces. Cell signalling can also result from signalling molecules that are integral membrane proteins on one cell binding receptors on an adjacent cell (Fig 13-3).