chapter 22 Cell division for growth and reproduction
DNA is the information store for most living organisms. In eukaryotes it is found in the nucleus in the form of linear chromosomes, composite structures made from DNA and specialised proteins, and in prokaryotes such as bacteria it is in the form of a circular chromosome. There are many differences in chromosome structure and cell division between prokaryotic and eukaryotic organisms, but this chapter will focus on the molecular mechanisms used in humans and other eukaryotic organisms. In an earlier chapter we saw that DNA is a very stable molecule because of its chemistry and structure, making it an ideal information storage molecule. For much of its life, an average cell normally only needs to check that the DNA has not been damaged, and does not need to synthesise more DNA. However, when that cell needs to undergo cell division to make new cells then the original cell must provide all the materials necessary for the new cells, including the DNA. For example, when skin is scratched to produce a small wound, new cells are required to heal the wound. Another example would be during growth and development where the body is getting larger and more cells are needed. There is one part of the body where another type of specialised cell division occurs. That is in the testes or ovaries where the production of sperm or eggs results from another type of cell division called meiosis.
Eukaryotic chromosomes
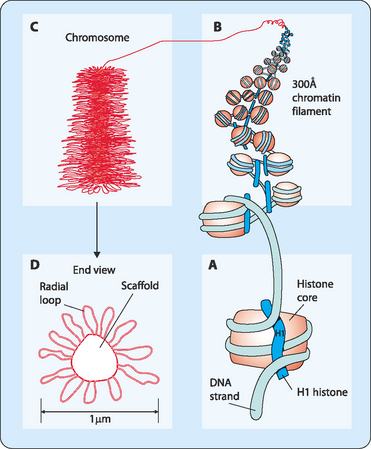
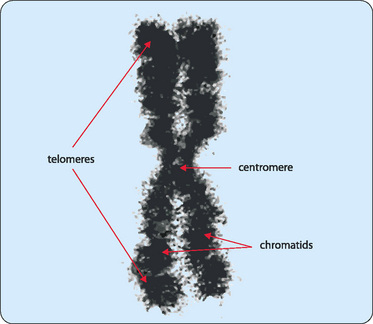
Chromosomes are not made of naked DNA but instead consist of chromatin, a complex of DNA and scaffolding proteins known as histones (Fig 22-1). These proteins form the structure on which the DNA can be wound, then coiled, and finally super-coiled. Initially the DNA in its double helical form is wound around core histones to form nucleosomes (diameter 10 nm). These structures look like a string of beads with some DNA tightly wound around a collection of histones then separated by a stretch of naked DNA before it winds around the next group of histones. Another specific protein, histone H1, is then added, which results in the nucleosomes associating closely to form a chromatin fibre (diameter 30 nm). More scaffolding proteins are then added and the fibre becomes super-coiled. Chromosomes vary in size and shape but each has several distinctive features (Fig 22-2). DNA replication produces pairs of identical sister chromatids, each of which generally carry the same genetic information. In metaphase, the chromatids interact at a point along their structure to form an X-shaped chromosome. This joining-point is referred to as the centromere. The position of the centromere can vary so the arms that are formed from a chromatid are not generally of equal length, though their position is always at the same point on each type of chromatid. Each of the arms of the chromosome carries a cap called a telomere, which protects the DNA strand from damage. These telomeres use a unique replication machinery that differs from the rest of the chromosome.
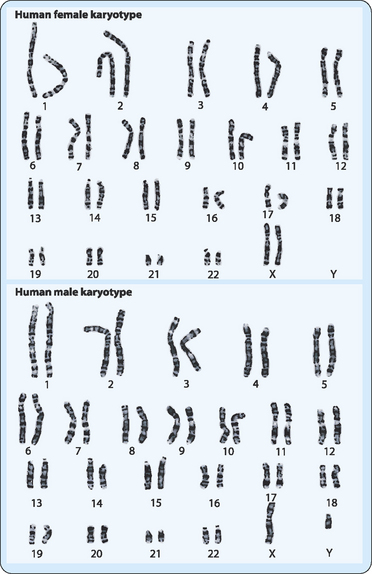
Different organisms have different numbers of chromosomes in their genetic composition (Table 22-1). The number can vary greatly and does not necessarily reflect the complexity of the organism. Thus, for example, a kingfisher has more than twice as many chromosomes as a human. The entire chromosome complement of an individual is referred to as a karyotype, which can be observed during mitotic metaphase, and is useful in the clinical setting. In most cases this is done by taking some cells from the individual (e.g. blood containing nucleated white cells), stimulating them with a drug to induce cell division, then processing the cells to see their chromosomes. A digital photograph of the chromosomes is then analysed by computer and the chromosomes sorted according to size and shape (Fig 22-3). The chromosomes can be stained to allow their finer details to be analysed. In humans, 44 of the 46 chromosomes in each cell can be sorted into 22 pairs of homologous chromosomes. Each of these pairs consist of chromosomes that are the same size, stain identically, have their centromeres in the same position, and carry information (i.e. the same genes) for the same inherited characters. However, the other two chromosomes cannot be placed in a matched pair because they are the ones which determine gender, the sex-determining chromosomes, of which there are two types, X and Y. A female has two X chromosomes whereas a male has one X and one Y chromosome. The other 22 pairs do not determine gender and are referred to as autosomes. These pairs of chromosomes are derived from the way in which we inherit genetic information from our parents (see below), with one of each pair of chromosomes originating from each parent.
TABLE 22-1 The number of chromosomes for different animals
| Animal | Number of chromosomes |
|---|---|
| Common fruit fly | 8 |
| Chinese hamster | 22 |
| Common earthworm | 36 |
| Laboratory mouse | 40 |
| Rabbit | 44 |
| Humans | 46 |
| Sheep | 54 |
| Dog | 78 |
| Cow | 60 |
| Horse | 64 |
| Goldfish | 100–104 |
| Kingfisher | 132 |
Note: The number of chromosomes required by each type of animal can vary greatly and does not seem to be related to its complexity.
The cell cycle
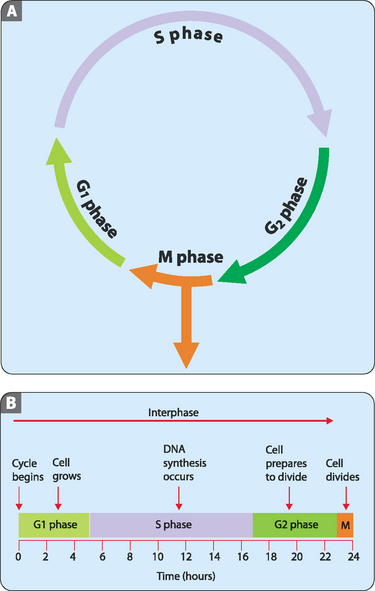
The vast majority of cells in the human body, which are referred to as somatic cells, are in the diploid state (2n). This means that they contain two complete genomes, in humans 46 chromosomes. Dividing cells exist in a series of defined states which together constitute the cell cycle (Fig 22-4A). For about 90% of the time most cells are in the stage referred to as interphase, during which the cell enlarges and copies its DNA in preparation for cell division. Interphase itself can be divided into three sub-stages: the G1 phase (1st gap), the S phase (synthesis), and the G2 phase (2nd gap). During all three of these sub-stages the production of new proteins and organelles proceeds but it is only in S phase that DNA synthesis, or replication, occurs. After completion of the interphase stage the cell undergoes mitosis, with the whole process from start to finish taking roughly 24 hours for rapidly dividing human cells (e.g. those of the skin, oral cavity, alimentary tract and vagina) (Fig 22-4B). However, a number of cells divide very slowly (e.g. those of the liver, pancreas and kidney) or not at all (e.g. cardiac and skeletal muscle or nervous tissue) over the course of a lifetime. These cells spend most of their lives in what is referred to as the G0, or resting phase.
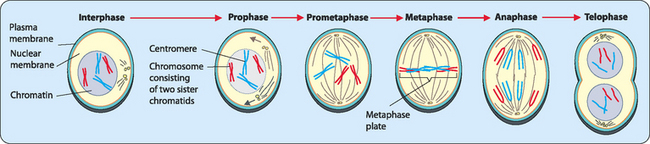
Mitosis—division for somatic cells
Mitosis consists of several discrete phases (Fig 22-5):
To generate two separate cells another process called cytokinesis
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


