15.2 Neuraxis310
15.3 Forebrain312
15.4 Brain stem318
15.5 Cranial nerve nuclei322
15.6 Vessels of the brain323
15.7 Motor systems and tracts325
15.8 Sensory systems and tracts328
15.9 Special sensation330
15.10 Clinical miscellany332
15.11 And finally…333
15.1. Brain vesicles, general terms, ventricles, CSF
Introduction
Introductory coverage of the brain considers its components from below upwards: hindbrain, midbrain, and forebrain. However, for looking at the brain in slightly more detail, beginning at section 15.2, it seems better to work down from the cerebral hemispheres to the medulla. Embryological terms are used both descriptively and clinically, so we begin with a brief developmental survey.
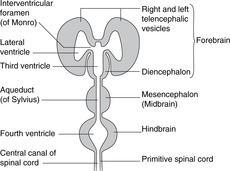
Hindbrain, midbrain, forebrain (Fig. 15.1)
Working up from the embryonic spinal cord, the three brain vesicles are the hindbrain, the midbrain and the forebrain.
 |
| Fig. 15.1 |
Hindbrain
This develops into the medulla, pons, and cerebellum. It contains the vital centres (cardiac, respiratory, etc.) and the nuclei for cranial nerves V–XII (facial sensation, and movements of the upper end of the gut tube for ingestion and phonation). The cerebellum is concerned with balance and position in space, receiving the majority of fibres from the vestibular apparatus, and with the coordination of complex patterns of skilled movement.
Midbrain
This part of the brain changes least during growth and maturation. It contains the nuclei for cranial nerves III and IV and the centres for eye reflexes (e.g. pupillary light reflex) and auditory reflexes.
Forebrain
This gives rise to the diencephalon (thalamic structures), and the right and left telencephalic derivatives – the cerebral hemispheres. The hemispheres are the only paired elements, all other components of the brain being unpaired. The cerebral hemispheres become very large and grow at first posteriorly, then downwards and forwards so that they are shaped like the horns of a ram, the inferior portions coming to lie on either side of the thalamus and midbrain, which are buried between them. The parts of the hemisphere are named according to the cranial bones to which they are related: the frontal, parietal, occipital and temporal lobes. These are not lobes in the sense of being clearly demarcated anatomical entities, but are simply areas.
• Diencephalic derivatives are concerned with motor and sensory coordination, regulation and control, and include the optic vesicles, which give rise to the optic nerves and retinas.
• Cerebral hemispheres are concerned with so-called ‘higher’ functions: intellectual, reasoning, long-term memory, conscious perception, and voluntary movement. In the cerebral hemispheres, unlike the spinal cord, grey matter is on the external surface, often called the cerebral cortex, or simply the cortex.
Brain stem
This term is used for the medulla, pons and midbrain together.
Terms derived from Greek:
• Hindbrain = rhombencephalon, rarely used.
• Midbrain = mesencephalon, widely used.
• Forebrain = prosencephalon, rarely used, of which:
– thalamic structures = diencephalon: widely used, and
– cerebral hemisphere = telencephalon: sometimes used.
15.2. Neuraxis (axis of the brain): motor and sensory areas
• In the medulla, pons and midbrain the ventral aspect of the neural tube is anterior, and the dorsal aspect is posterior, much as in the spinal cord. Above the midbrain, the axis bends forwards so that in the diencephalon, the ventral aspect is inferior and the dorsal surface superior.
• In the spinal cord, the general rule is that dorsal components tend to be concerned with afferent (sensory) functions, and ventral with efferent (motor) functions. In the medulla it is as if the two dorsal (sensory) components have been dragged laterally so that sensory components are no longer dorsal but lateral, and motor components now not so much anterior as medial. Higher in the brain stem the pattern becomes less and less obvious, and it has all but disappeared in the diencephalon.
Nucleus
A nucleus is a collection in the central nervous system of neuronal cell bodies serving similar functions. A collection of cell bodies in the peripheral nervous system is a ganglion – except that some groups of cell bodies in the brain are known as the basal ganglia, a term sanctified by long-standing use.
Ventricular system, cerebrospinal fluid
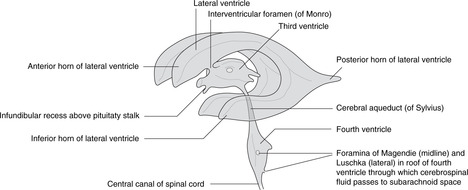
Ventricles (Figs 15.1, 15.2)
Dilatations of the central canal of the neural tube form the cerebral ventricles.
• Hindbrain. The central canal forms the fourth ventricle, continuous below with the central canal of the spinal cord. The ventral surface of the ventricle is the diamond-shaped floor (diamond-shaped brain: rhombencephalon). The thin dorsal surface is the roof and has three openings into the subarachnoid space: one median aperture, the foramen of Magendie, and two lateral apertures, the foramina of Luschka.
• Midbrain. The central canal forms the cerebral aqueduct of Sylvius.
• Diencephalon. The central cavity of the diencephalon forms the third ventricle. This is narrow from side to side (1 or 2 mm) but 10–15 mm in height. Its floor is the hypothalamus and its lateral walls the thalamus of each side, which may touch each other in the midline giving the interthalamic adhesion. You can regard the diencephalon as the structures surrounding the third ventricle or attached to its walls, thus including the optic chiasma, nerve and retina, and the posterior pituitary.
• Telencephalon. The cavities of the cerebral hemispheres are the right and left lateral ventricles. They are adjacent to the midline and are separated from each other by the septum pellucidum, a thin sheet of tissue of no neurological significance. Each lateral ventricle communicates with the third ventricle through an interventricular foramen of Monro, but the two lateral ventricles do not communicate with each other. The lateral ventricle assumes the shape of the hemisphere in which it is situated: there is an anterior (frontal) horn, a posterior (occipital) horn, and an inferior (temporal) horn.
Cerebrospinal fluid
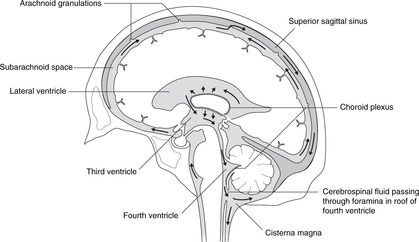
Cerebrospinal fluid is produced by the choroid plexus, a vascular plexus present in all ventricles formed where vessels of the pia mater (external) come into contact with the ependymal lining of the central canal (i.e. there is no intervening neural tissue).
Circulation of cerebrospinal fluid (Fig. 15.3)
From the lateral ventricles, cerebrospinal fluid flows through the foramina of Monro to the third ventricle. It continues through the cerebral aqueduct to the fourth ventricle where some may enter the central canal of the spinal cord, but most passes to the subarachnoid space through foramina in the roof. It flows around the brain and spinal cord and is absorbed back into the venous system at the arachnoid granulations (see section 14.12, p. 297, Fig. 14.37).
 |
| Fig. 15.3 |
Hydrocephalus may be caused by overproduction of cerebrospinal fluid, or reduced absorption (e.g. thrombosis of the superior sagittal sinus). It may be generalised or local, for example blockage of the cerebral aqueduct would cause hydrocephalus in the lateral and third ventricles, but not in the fourth or in the subarachnoid space.
Tracts in the central nervous system, internal capsule
The brain stem contains tracts: bundles of myelinated axons (i.e. white matter) all serving similar functions. They can be divided into:
• motor or descending tracts which convey impulses away from the cerebral hemispheres; and
• sensory or ascending tracts which convey impulses towards the hemispheres.
Each of these may be further subdivided into named tracts connecting specific areas, the name of the tract generally being derived from the origin and the destination. Thus, the corticospinal tract conducts impulses from the cortex to the spinal cord – a motor tract; the spinothalamic tract conducts impulses from the spinal cord to the thalamus – a sensory tract.
Internal capsule
In the region where telencephalic and diencephalic regions meet, all the tracts in and out of the cerebral hemispheres are within a fairly small area on each side forming a sheet of white matter. This is the internal capsule, and it is covered in more detail later.
15.3. Forebrain
The cerebral cortex on each side is shaped like a ram’s horn. Its parts are named according to the bones they are related to. The principal functional areas are motor, sensory, auditory, and visual. The limbic system is the area responsible for emotions, short-term memory and instinctual behaviour. The central part of the brain, on either side of the third ventricle, is the diencephalon consisting mainly of the thalamus and hypothalamus. All tracts conveying impulses between cerebral cortex and brain stem pass in the internal capsule, which lies lateral to the thalamus, between it and the cerebral cortex. Lesions here are usually catastrophic because so many axons are packed into such a small area. The basal ganglia are a group of nuclei adjacent to the diencephalon, and in the midbrain.
You should:
• list the principal parts of the cortex
• identify the site of the motor, sensory, auditory and visual areas
• list the components of the limbic system and their functions, in as far as they are known
• list the components of the basal ganglia and their functions, in as far as they are known
• describe the anatomy of the internal capsule, and the term ‘stroke’.
Cortex
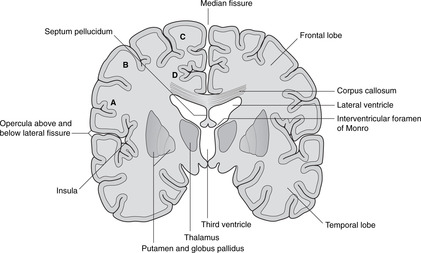
The cerebral cortex is shown in Figures 15.4 and 15.5. The numerous small fissures are sulci (singular: sulcus) and the raised areas between them are gyri (singular: gyrus).
• The large fissure between the parietal and temporal lobes is the lateral fissure and its edges are the opercula (singular: operculum). If you separate the opercula you will see a buried area of cortex deep in the lateral fissure: this is the insula.
• Commissures. Numerous bundles of fibres (white matter) connect the two sides: these are the commissures, and the largest, deep in the sagittal fissure, is the corpus callosum. The anterior portion of the corpus callosum is the genu, the central part the body, and the posterior part the splenium. There are other commissures: the anterior commissure is part of the anterior wall of the third ventricle, and the posterior and habenular commissures are in the posterior wall of the third ventricle.
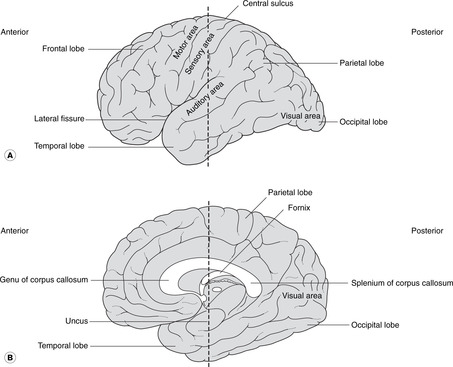 |
| Fig. 15.4 The cerebral hemispheres: (A) lateral aspect; (B) medial aspect. The dotted line indicates the plane of the section in Figure. 15.5. |
Functional areas of cortex (Figs 15.4, 15.5)
Motor, sensory
On the lateral aspect of the cortex is the central sulcus (it is neither central nor easily identifiable), the approximate surface marking of its inferior limit being the pterion. The cortex anterior to the central sulcus, the precentral gyrus, is the main motor area concerned with the initiation of voluntary movement. The cortex posterior to it, the postcentral gyrus, is the main sensory area.
• For both the motor and sensory cortex the inferior (lateral) portion is concerned with the head. The rest of the body is represented sequentially passing superiorly and medially over the cortex so that the cortical areas for the lower limbs are on the medial aspect of each cerebral hemisphere with the foot area being adjacent to the corpus callosum.
• It is a fact, although one without convincing explanation, that the right cortex serves the left side of the body and vice versa.
• Broca’s area is adjacent to the head area of the dominant motor cortex (usually the left in a right-handed person). It is in the frontal lobe, immediately above the lateral fissure, and is concerned with the motor mechanisms of speech. Damage to it (for example occlusion of a branch of the middle cerebral artery) leads to motor speech aphasia (aphasia: wordless): it is not that the patient does not know the word, it is that he can not muster the motor forces to enunciate it.
Visual, auditory, olfactory
• The visual cortex is in the occipital lobe (posterior)
• The auditory cortex is on the superior surface of the temporal lobe.
• The olfactory cortex is on the under surface of the temporal lobe, principally the uncus.
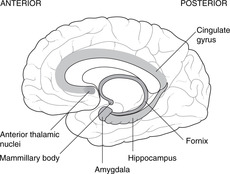
Limbic system (Fig. 15.6)
The Latin word limbus means margin or border. The limbic system develops from the border of the telencephalon adjacent to the diencephalon. Like many other telencephalic derivatives, this assumes the ram’s-horn shape. Components of the limbic system include the fornix, the hippocampus (medial to the inferior horn of the lateral ventricle), the amygdala, the uncus and the cingulate gyrus (medially, just above the corpus callosum). The limbic system has connections with olfactory and taste fibres, and with certain diencephalic structures, e.g. the mammillary bodies and anterior thalamic nuclei. Limbic functions include elements of emotion, recent memory, olfaction, taste, sexual urges, and basic instinctive drives: smells and tastes can evoke memories, emotions, mood changes and sexual responses.
Destruction or degeneration of the mammillary bodies
This occurs in alcoholics and as a result of vitamin B1 (thiamine) deficiency. Short-term memory is lost, so patients tend to cover up this defect by inventing fiction: confabulation (Korsakov’s syndrome). It has even been known to occur in operations for pituitary tumours that have gone wrong.
Congenital absence of the amygdala
Patients without an amygdala, and such people are rarely found, neither feel rage, nor recognise it in others – a blissful state, one would think. They may also exhibit hypersexuality. It is difficult to think of the amygdala as anything other than the organ of guilt and repression.
Dementias, Alzheimer’s disease
Progressive dementias may arise from disorders of the hippocampus and its connections, associated with a loss of cholinergic neurons in Meynert’s nucleus in the anterior limbic system.
 |
| Fig. 15.6 |
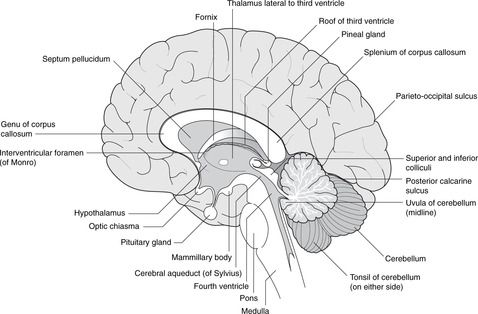
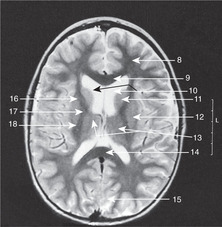
Diencephalon (Figs 15.7, 15.8, 15.10)
• The hypothalamus forms the floor of the third ventricle. Its most obvious features when inspecting the brain from below are the pituitary stalk and the mammillary bodies (mammilla = little mamma or breast). The hypothalamus contains the centres regulating appetite and much autonomic activity (e.g. temperature regulation) as well as controlling the activity of the pituitary gland.
– The posterior pituitary is a component part of the hypothalamus with direct neural connections from the supraoptic and paraventricular nuclei of the hypothalamus. Hormones released by the posterior pituitary (oxytocin, vasopressin) are manufactured in the cell bodies of neurons in these nuclei, and travel down axons to the posterior pituitary where they are released into the bloodstream.
– The anterior pituitary is a separate gland that migrates from the roof of the primitive pharynx. The hypothalamus controls it by releasing hormones into the hypophyseal portal system of veins, through which they are transported to the anterior pituitary to act upon its component cells.
• The roof of the third ventricle contains no neural tissue except for commissural fibres which arise anteriorly and grow backwards, over the roof of the third ventricle, to form the corpus callosum.
• The epithalamus is the posterior region of the third ventricle: it contains the pineal gland.
• The metathalamus, the most lateral part of the diencephalon, consists of the geniculate bodies (lateral: part of the visual pathway; medial: part of the auditory pathway).
Pituitary tumours
Immediately anterior to the pituitary stalk is the optic chiasma, which may be damaged by enlarging pituitary tumours causing a bitemporal hemianopia (see section 14.11, pp. 293, 294).
Pineal
The pineal gland often calcifies with advancing age. The advantage of this is that it provides a landmark for the assessment of whether or not there is any displacement of the brain: the pineal gland is a midline structure.
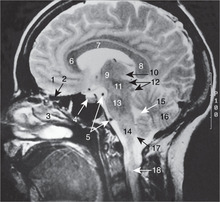
Internal capsule, basal ganglia (Figs 15.5, Fig. 15.10, Fig. 15.11, Fig. 15.12 and Fig. 15.13)
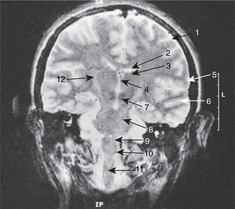
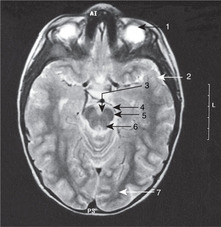
Internal capsule
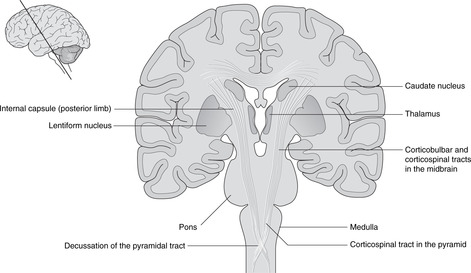
The relations of the internal capsule are:
• medial to the posterior limb: the thalamus
• medial to the anterior limb: the caudate nucleus, another of the structures stretched into the shape of a ram’s horn by the growth of the brain
• lateral to both limbs: the lentiform nucleus, itself composed of a medial globus pallidus and a lateral putamen.
There are numerous fibres crossing the internal capsule linking the thalamus, caudate and lentiform nuclei, and when a fresh or fixed brain is cut, these are evident as striations. For this reason, this area was originally called the corpus striatum, and this term, although of doubtful validity, is still used.
Basal ganglia
The caudate nucleus, putamen and globus pallidus, together with the substantia nigra in the midbrain (see below) and other nuclei, constitute the basal ganglia. Their function is the control of movement, and disease of them leads to unwanted involuntary movements, for example tremors, writhing (athetosis), fidgeting (chorea) and uncontrolled explosive movements (hemiballismus). Parkinson’s disease results from disorders of the substantia nigra, and chorea and athetosis from disorders of the caudate and lentiform nuclei.
Lateral to the basal ganglia
Lateral to the basal ganglia are, in order, the external capsule, the claustrum (grey matter), the extreme capsule and the insular cortex. None of these areas is very important clinically.
Stroke
When disease (such as haemorrhage or arterial occlusion) affects the internal capsule, the result is a profound and often catastrophic loss of function because so many fibres, both motor and sensory, are concentrated into such a small area. This is, in common parlance, a stroke and is discussed in more detail later.
15.4. Brain stem
The brain stem consists of grey matter made up of cell bodies grouped as nuclei, and white matter: the myelinated axons of fibre tracts. Nuclei of the cranial nerves are prominent in the brain stem. Those of III and IV are in the midbrain, those of V–XII in the pons and medulla. Some of the tracts are short local connections which run in all directions between nuclei, and others are long motor and sensory tracts conveying information between cerebral hemispheres and spinal cord. The midbrain houses nuclei concerned with visual and auditory reflexes. The cerebellum is posterior to the pons and receives most of the information from the vestibular system. Cerebellar lesions are ipsilateral, unlike those of the rest of the brain which are usually contralateral.
Summary of brain stem nuclei
• Motor nuclei of cranial nerves III–XII.
• Parasympathetic (visceral motor) nuclei: Edinger–Westphal, superior salivatory, inferior salivatory, dorsal motor nucleus of the vagus (Fig. 14.14, p. 258).
• Sensory nuclei: trigeminal sensory nuclei for cutaneous and oral sensation, nucleus of the solitary tract for visceral sensation (e.g. taste).
• Other named nuclei for visual and auditory reflexes, as outlined in the text below.
• Nuclei of the reticular formation concerned with vital centres and arousal. These are hard to pinpoint accurately and for clinical purposes it is unnecessary to do so. One definition of the reticular formation is that it is what is left when you have removed everything else with a specific name.
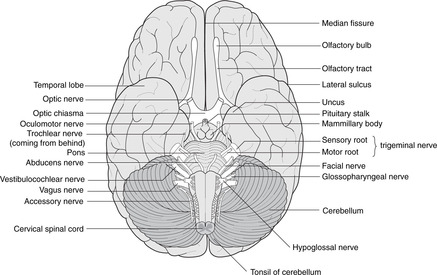
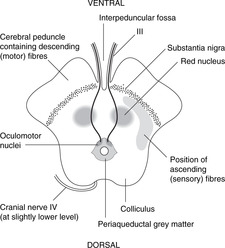
Midbrain (Fig. 15.14)
The midbrain passes through the tentorial notch of the tentorium cerebelli. It has a very distinctive shape in cross-section and is easy to draw. This is worth the effort since the midbrain is an important area from the point of view of pupillary and accommodation reflexes, descending motor tracts and other clinical conditions. Its parts are as follows.
• Tectum. This is the area dorsal to the aqueduct. There are four dorsal prominences, two superior colliculi and two inferior colliculi (also known together as the corpora quadrigemina). The superior colliculi are associated with visual reflexes and the inferior colliculi with auditory reflexes. Cranial nerve IV is attached between them.
• Periaqueductal (around the aqueduct) grey matter. This contains the nuclei of cranial nerves III and IV and the medial longitudinal fasciculus, which integrates the nuclei of III, IV and VI with vestibular information, amongst other things. In this area, fibres involved in the pupillary light reflex pass from the optic tracts to the Edinger–Westphal nucleus of III.
• The tegmentum is the area ventral to the aqueduct. One of its components is the red nucleus, to all intents and purposes an outpost of the cerebellum (see below).
• Substantia nigra (Latin: black substance). This is a dark area in the ventral tegmentum. The pigment is melanin, a reminder of the amine biochemistry commonly found in derivatives of neuroectoderm. Depleted levels of dopamine in this area lead to symptoms of Parkinson’s disease (e.g. resting tremor): functionally, this is one of the basal ganglia.
• Anterior to the substantia nigra are two large projections facing slightly laterally. These contain the main descending motor fibres called, variously, the cerebral peduncles, basis pedunculi, or cerebral crura, each being the crus cerebri. This text uses the first term.
• The fossa between the peduncles is the interpeduncular fossa in which cranial nerve III is attached.
• The long sensory tracts conveying information from the spinal cord and the lower brain stem to the thalamus are just behind the substantia nigra, extending posteriorly on to the lateral aspect of the midbrain. The long motor tracts are in the cerebral peduncles. (Dorsal: sensory; ventral: motor.)
Pons (Figs 15.7, 15.8)
The pons is that part of the brain stem to which the cerebellum and cranial nerve V are attached. It contains scattered nuclei concerned with cerebellar connections, and nuclei of cranial nerves V, VI and VII, although the course of fibres from VI and VII means that these nerves are attached lower down the brain stem at the pontomedullary junction. For reasons which are not clear, fibres of VII pass at first dorsally before looping round the nucleus of VI and turning anteriorly.
The locus coeruleus (Latin: blue place) is a pigmented area (amines again) in the dorsal pons. It is concerned with the limbic system and autonomic functions and is of interest to neuroscientists as a focus for drug action.
The long motor and sensory tracts are scattered in numerous bundles as they pass through the pontine substance.
Cerebellum (Figs 15.7, 15.8)
The cerebellum is an outgrowth of the dorsal pons, separated from it by the fourth ventricle. On each side, the white matter connecting the pons and cerebellum forms the superior, middle and inferior cerebellar peduncles. Despite all the work of neuroscientists, the cerebellum remains somewhat mysterious: it is concerned with the coordination of motor activity, the regulation of muscle tone and the maintenance of equilibrium.
Cerebellar cortex
The cerebellar cortex has superficial grey matter, inside which is white matter and, most deeply of all, the four paired cerebellar nuclei. Cerebellar equivalents of the cerebral gyri are folia (thinner and straighter than gyri). The midline portion is the vermis, the most inferior part of which is the uvula and the most superior part (on the roof of the fourth ventricle) the lingula. Laterally, the most inferior parts of the cerebellar hemispheres are the tonsils, so, as at the back of the mouth, the uvula is between the tonsils.
Vestibulocerebellum
The cerebellum, more obviously than other parts of the nervous system, displays its evolution. The earliest part to arise was that concerned with vestibular impulses, the vestibulocerebellum, and it is largely separate from the rest of the cerebellum, although this is not evident to the naked eye. It is the flocculonodular node, the floccules being on either side joined in the midline at the nodule. The deep cerebellar nucleus of the vestibulocerebellum is the fastigial nucleus, which has connections with:
• cranial nerve VIII: most fibres from the vestibular apparatus pass to the cerebellum through the inferior cerebellar peduncle
• the medial longitudinal fasciculus for the coordination of vestibular impulses and eye movements (nuclei of III, IV, VI).
Other parts of the cerebellum. Subsequently (in evolutionary terms), parts of the cerebellum acquired connections with the spinal cord (for movements of the limbs and trunk) and most recently with the cerebral cortex. The deep cerebellar nuclei for spinal connections are the globose and emboliform, and for cerebral connections the dentate nucleus.
• Impulses from the spinal cord arrive in spinocerebellar tracts.
• Impulses to and from the cerebral cortex pass from the dentate nucleus through the superior cerebellar peduncle to the red nucleus (midbrain) and on to the cerebral cortex (dentato-rubro-thalamic tract). Connections from the red nucleus pass to and from the spinal cord (thus rubrospinal, spinorubral) for further integrative activity.
Cerebellar disease
Disease of the cerebellum causes ipsilateral symptoms, not contralateral, manifested principally as a lack of coordination called ataxia. This gives rise to an intention tremor; that is to say there is no tremor present at rest, but as soon as any movement is attempted, lack of coordination becomes apparent. This contrasts with the tremor of basal ganglia disease, which is present at rest, but which disappears when purposeful movement is attempted. The resultant speech patterns are called scanning speech (it is as if the person is scanning his vocabulary for the right word, so laboured is the progress); the lack of coordination of eye movement is nystagmus. Cerebellar lesions are perfectly mimicked by drunkenness, which is nothing more or less than cerebellar poisoning by alcohol, and it is not unknown for a person with cerebellar disease to be arrested on suspicion of being intoxicated.
Pontomedullary junction (Fig. 15.15)
The pontomedullary junction is marked anteriorly by a transverse sulcus where VI is attached medially, and VII and VIII laterally in the cerebellopontine angle.
 |
| Fig. 15.14 |
Medulla (Fig. 15.15)
Anterior aspect. Four swellings are visible, two on either side of the midline.
• The medial swelling on each side is the pyramid containing the descending motor corticospinal tracts to the opposite side of the body. The rootlets of XII are attached laterally to these swellings.
• Lateral to the pyramid is the olive containing the inferior olivary nucleus, a convoluted band of grey matter which, like the red nucleus in the midbrain, is an outpost of the cerebellum. Lateral to the olive are the rootlets of, from above down, IX, X and XI.
Posterior aspect.
Two tubercles are visible on each side, the gracile medially and the cuneate laterally. These are concerned with sensation from the spinal cord (see below).
Nuclei
The hypoglossal nucleus is close to the midline and the floor of the fourth ventricle. The motor nuclei of the vagus nerve are lateral to this (dorsal motor and nucleus ambiguus), and the ‘spinal’ part of the trigeminal sensory nucleus more lateral still.
Medulla/spinal cord junction
This is not at the foramen magnum, but a few millimetres below. It means that C1 and C2 nerves pass slightly upwards from their spinal cord origin to emerge from the vertebral column.
Some features of the fourth ventricle
• In the floor, the facial colliculus is a small swelling lateral to the central region of the ventral median sulcus. It is caused by the internal genu of the fibres of VII looping around the nucleus of VI. Lower down is the hypoglossal trigone, immediately over the nucleus of XII.
• In the roof, the foramina of Luschka (right and left) and Magendie (median) allow cerebrospinal fluid to pass from the ventricular system to the subarachnoid space.
15.5. Cranial nerve nuclei
Cranial nerve nuclei can be grouped by functional type: somatic motor, branchiomotor, visceral motor (parasympathetic), somatic sensory, visceral sensory and special sensory. These groups are disposed vertically in the brain stem, somatic motor being the most medial, with sensory nuclei more laterally sited. Not all nuclei contribute to all cranial nerves, some nerves containing only somatic motor fibres, and some only sensory fibres. Mixed nerves are those that supply the upper end of the gut tube – in embryological terms, these are the nerves of the branchial arches.
Brain stem nuclei
Lesions of the brain stem give rise to symptoms according to which cranial nerves arise near the lesion. Motor impulses in cranial nerves arise from brain stem motor nuclei, and sensory impulses in cranial nerves pass to brain stem nuclei. Table 15.1, Table 15.2 and Table 15.3 set out these nuclei in detail. As a simplification, it is enough for you to realise that:
• midbrain lesions cause disturbances of eye movements
• pontine lesions cause disturbances of mandibular movement
• medullary lesions cause disturbances of phonation and swallowing.
| Nucleus | Position | Nerve | Function |
|---|---|---|---|
| Oculomotor | Upper midbrain | Oculomotor III | Eyeball movements |
| Trochlear | Lower midbrain | Trochlear IV | Eyeball movements |
| Trigeminal | Pons | Mandibular Vc | Mastication |
| Abducens | Pons | Abducens VI | Eyeball movements |
| Facial | Pons | Facial VII | Facial expression, etc., stapedius |
| Nucleus ambiguus | Medulla | Glossopharyngeal IX | Swallowing, phonation |
| Vagus X | |||
| Accessory | See text | Accessory XI | Sternocleidomastoid, trapezius |
| Hypoglossal | Medulla | Hypoglossal XII | Tongue |
| Nucleus | Position | Nerve(s) | Function |
|---|---|---|---|
| Edinger–Westphal | Midbrain | Oculomotor III | Pupilloconstriction, accommodation |
| Superior salivatory | Pons | Facial VII, Vb | Secretomotor: nasal, palatine, lacrimal glands |
| Superior salivatory | Pons | Facial VII,Vc | Secretomotor: submandibular, sublingual glands |
| Inferior salivatory | Upper medulla |



