Antimicrobial Resistance and Healthcare-Associated Infections
Antimicrobial Resistance and Healthcare-Associated Infections
Gary L. French
HOSPITAL PATHOGENS TEND TO BE ANTIMICROBIAL RESISTANT AND MULTIPLY RESISTANT
To be successful as hospital pathogens, healthcare-associated bacteria must be able to establish themselves and survive in the hospital environment, colonize the mucosa and skin of patients and staff members, survive on various surfaces during patient-to-patient transmission, and resist antibiotic and sometimes antiseptic therapy (1). Inherent multiple antibiotic resistance, and the ability to acquire additional genetic resistance factors in the face of increasing use of antibiotics, are important for survival. Numerous reports show that microorganisms causing healthcare-associated infection and colonizing patients and healthcare workers are more antibiotic-resistant than those in the community and that, within the hospital, resistance rates are high in units (such as intensive care, hematology and oncology, and renal and liver units) where antimicrobial use is highest. Indeed, risk factors for colonization and infection with multidrug-resistant (MDR) pathogens include prolonged hospital stay, prior antimicrobial therapy, and admission to intensive care units (ICUs).
EMERGENCE OF ANTIBIOTIC RESISTANCE IN HEALTHCARE-ASSOCIATED PATHOGENS
Innate and Acquired Antimicrobial Resistance
In his initial studies in 1929, Fleming noted that penicillin was highly active against some microorganisms, especially the gram-positive staphylococci and streptococci, but inactive against others, especially the gram-negative coliforms (2). This phenomenon of innate susceptibility or resistance to different agents among different bacterial species continued to be seen with broad-spectrum antimicrobials. For example, Klebsiella pneumoniae is usually resistant to ampicillin; Enterobacter spp. to ampicillin and many cephalosporins; enterococci to cephalosporins and quinolones; and Pseudomonas aeruginosa to pencilin, ampicillin, cephalosporins, and other groups. The β-hemolytic streptococci remain susceptible to penicillin and the anaerobes to metronidazole, but increasing numbers of isolates of many other species have acquired resistance to agents to which they were initially innately susceptible.
Free-living environmental bacteria are of low virulence for humans but are often inherently resistant to common antimicrobials. This is probably because they are adapted to live in soil and water where they are exposed to naturally occurring antimicrobial substances. Environmental species with inherent antimicrobial resistance include various Pseudomonas spp., Acinetobacter, Burkholderia, Stenotrophomonas, and Ralstonia. Although they rarely infect healthy individuals, these microorganisms may contaminate hospital environments and equipment and then colonize and infect compromised patients, producing antibiotic-resistant opportunistic infection.
Naturally sensitive bacteria may acquire antibiotic resistance caused by a number of mechanisms, with the most common probably being the production of drugdestroying enzymes (3,4). This is the typical mechanism by which microorganisms such as Staphylococcus aureus and Escherichia coli and other gram-negative bacteria acquire resistance to ampicillin, aminoglycosides, and chloramphenicol. There may be alterations in the permeability of the cell wall, preventing antibiotics from reaching their target sites, or there may be increased antibiotic efflux, resulting in the same effect. This is the common mechanism of tetracycline resistance and is one of the ways in which microorganisms such as P. aeruginosa may acquire broadrange resistance to several aminoglycosides and other agents simultaneously. Alterations in target sites prevent antibiotics from binding to their sites of action. Changes in the affinities of penicillin-binding proteins result in methicillin resistance in staphylococci, penicillin resistance in pneumococci, and ampicillin resistance in enterococci. Alterations in ribosomal-binding sites may produce acquired resistance to rifampin, fusidic acid and the macrolides, and alteration of DNA gyrase is the common mechanism of quinolone resistance. Alterations (or substitutions) of enzymes in metabolic pathways are responsible for resistance to sulfonamides and trimethoprim that block bacterial folate metabolism.
Acquired resistance may emerge by genetic mutation, which occurs relatively frequently in rapidly multiplying microorganisms or by acquisition of resistance genes from other bacteria. The horizontal spread of resistance genes among bacteria by plasmid transfer is sometimes called “infectious resistance.” The transmission of DNA between bacteria may occur by bacteriophage transduction (as in the transmission of penicillinase-mediated penicillin-resistance in S. aureus), conjugation (the common mechanism of transfer between gram-negative species), or transformation. Transformation was previously regarded as a relatively unimportant mechanism of resistance transfer in clinical bacteria, but there is increasing evidence for its importance in the emergence of resistance in gram-positive microorganisms. Although the host range of many plasmids is restricted and gram-positive and gramnegative microorganisms tend not to share resistance genes, plasmids can be exchanged between different bacterial species. For example, most ampicillin resistance in Haemophilus influenzae is mediated by a β-lactamase that probably originated from E. coli.
Resistance genes may be encoded on a variety of transferable elements, including transposons and integrons that can insert into both chromosomes and plasmids. The combination of several insertion elements may create large multiple resistance-gene packages (5). Integrons encoding multiple antimicrobial resistances are now widespread in Enterobacteriaceae in both hospitals and the community (6,7). There is continuous horizontal transfer of these resistance genes between and within species, and acquisition of multiple resistances favors the proliferation of certain cross-infecting microorganisms in hospitals (8). Some species of both good and opportunistic pathogens appear to have a special ability to accumulate multiple resistance genes and become increasingly MDR. Examples of successful MDR healthcare-associated pathogens include S. aureus and coagulase-negative staphylococci (especially methicillinresistant strains), Enterococcus faecium (especially glycopeptide-resistant strains), K. pneumoniae, Acinetobacter baumannii, and P. aeruginosa.
Antibiotic Use and Antibiotic Resistance
Hospital patients often have compromised host defenses due to treatment or underlying disease and are therefore at risk of acquiring infection with both virulent and opportunistic pathogens. Since antibiotic use is concentrated in hospitals, both types of pathogen are more likely to survive and proliferate in the hospital environment and colonize patients if they are resistant to common antimicrobials. Antibiotic therapy tends to suppress innately sensitive commensal bacteria and encourage their replacement with resistant microorganisms. Initially, the more resistant members of generally sensitive species are selected (a shift within a species to a population with increased resistance), and then the inherently resistant genera emerge (a shift to colonization with more resistant species). Patients who receive multiple courses of antibiotics commonly become colonized by increasingly resistant microorganisms, often suffering sequential infections with bacteria such as Klebsiella, P. aeruginosa, enterococci and Acinetobacter, and finally with antibacterial resistant fungi such as Candida spp. The tendency for antibiotic use to promote the emergence of resistant pathogens is called “antibiotic pressure” and is an inevitable evolutionary process. Despite methodological difficulties, there are many reports of resistance rising during increased antibiotic use and falling after a reduction in use (9).
A change in the pattern of serious healthcare-associated infection after the introduction of antibiotics was first noted by Finland and his colleagues in 1959 (10). Between 1935 and 1957, antibiotic-sensitive gram-positive pathogens were replaced by penicillin-resistant S. aureus and multi-resistant gram-negative bacteria such as E. coli, Klebsiella, and Proteus spp. Once the emergence of resistant opportunistic pathogens had been recognized, new, more effective drugs were developed for their treatment. The worldwide problem of the multiresistant “hospital staphylococcus” in the 1960s diminished after the introduction of methicillin and other penicillinase-stable penicillins (11); outbreaks of gentamicin-resistant Klebsiella and other gram-negative microorganisms seen in the 1970s waned in the 1980s with the use of newer aminoglycosides and cephalosporins.
After the 1980s, the pattern changed again with a dramatic increase in healthcare-associated infections with multiply resistant gram-positive bacteria (1,12). Methicillin-resistant S. aureus (MRSA), resistant to all β-lactams and to many other previously effective agents, has emerged as a worldwide cause of healthcare-associated infections and outbreaks associated with serious morbidity and mortality (13, 14and15). Coagulase-negative staphylococci are increasingly common healthcare-associated pathogens, partly because they too are often resistant to methicillin and other agents but also because many strains produce an extracellular slime (16,17) that enables them to colonize the intravascular and other plastic prostheses that are increasingly used in modern medicine. Finally, many antibiotics used for gram-negative healthcare-associated infections, including ampicillin, the aminoglycosides, cephalosporins and quinolones, are ineffective against enterococci, which have also emerged as important causes of healthcare-associated infection (18).
This is, of course, a continuing dynamic situation. Resistant gram-positive bacteria remain a major feature of healthcare-associated infection but MDR gram-negative bacteria continue as important healthcare-associated pathogens, especially with the emergence of extendedspectrum β-lactamase (ESBL)-producing and multiply resistant strains of K. pneumoniae, Enterobacter, Serratia, and, most recently, E. coli (19). Broad-spectrum antibiotic use encourages the proliferation of antibiotic-resistant and toxigenic strains of Clostridium difficile in the bowel that cause diarrhea and pseudomembranous colitis. There has recently been an increase in community and healthcareassociated infections with C. difficile associated with the worldwide dissemination of highly virulent strains (20). Thus, healthcare-associated infection is now microbiologically heterogeneous, often being caused simultaneously by several different species of MDR gram-positive and gram-negative bacteria. Table 86-1 shows the distribution of healthcare-associated pathogens reported in the United States in 2003 (21).
Table 86-2 shows the rates of antibiotic resistance in bacterial isolates from hospitals taking part in the U.S. National Nosocomial Infections Surveillance (NNIS) system from 1992 to 2004 (22). Antibiotic resistance rates were highest in ICUs, followed by non-ICU wards, and then outpatients, reflecting the levels of compromise, antimicrobial courses, and length of hospitalization of the patients in these three areas. ICUs had especially high rates of methicillin resistance in both coagulase-negative and coagulase-positive staphylococci, high rates of quinolone and carbapenem resistance in P. aeruginosa, and high rates of resistance to third-generation cephalosporins in Enterobacter spp.
TABLE 86-1 Percentage of Bacterial Isolates Associated with Healthcare-Associated Infection of Different Types in Intensive Care Units Reported to the National Nosocomial Infections Surveillance System, 2003
Pneumo
BSI
SSI
UTI
Pathogen
(n = 4,365)
(n = 2,351)
(n = 2,984)
(n = 4,109)
Gram-negative
Escherichia coli
5.0
3.3
6.5
26.0
Klebsiella pneumonia
7.2
4.2
3.0
9.8
Enterobacter spp.
10.0
4.4
9.0
6.9
Serratia marcescens
4.7
2.3
2.0
1.6
Pseudomonas spp.
18.1
3.4
9.5
16.3
Acinetobacter spp.
6.9
2.4
2.1
1.6
Other
14.1
3.8
9.8
10.7
Gram-positive
Coagulase-negative staphylococci
1.8
42.9
15.9
4.9
Staphylococcus aureus
27.8
14.3
22.5
3.6
Enterococci
1.3
14.5
13.9
17.4
Other
3.2
4.5
5.8
1.2
Pneumo, pneumonia; BSI, bloodstream infection; SSI, surgical site infection; UTI, urinary tract infection. (From Gaynes R, Edwards JR; the National Nosocomial Infections Surveillance System. Overview of nosocomial infections caused by gram-negative bacilli. Clin Infect Dis 2005;41:848-854.)
Table 86-3 shows results from the NNIS system comparing resistance rates in US ICUs for various pathogens isolated in 1994 to 1998 and 1999 (22,23). During this period, there was a dramatic increase in resistance rates for several important healthcare-associated pathogens. In particular, there was a 47% increase in vancomycin resistance in enterococci, a 43% increase in methicillin resistance in S. aureus, and an increase in resistance in P. aeruginosa of 35% and 49% to imipenem and the quinolones, respectively.
The European Antimicrobial Resistance Surveillance System (EARSS) has been reporting on antimicrobial resistance rates in invasive (mainly bloodstream) isolates from European countries since 1999. In 2008, the EARSS network comprised almost 900 microbiological laboratories serving more than 1,500 hospitals in 33 countries and provided susceptibility data on more than 700,000 invasive isolates (24). The annual reports from this organization give good data on European trends and can be used to illustrate the generally growing problem of antimicrobial resistance in clinically important bacteria while emphasizing significant regional and geographical differences. EARSS data on resistance trends are discussed under the individual pathogens, later in this chapter.
THE CLINICAL AND ECONOMIC IMPACT OF ANTIMICROBIAL RESISTANCE
Increasing antimicrobial resistance and multiple resistance results in increasing difficulties in the treatment of bacterial infections. Resistance leads to inappropriate empirical therapy; delay in starting effective treatment; and the use of less effective, more toxic, and more expensive drugs (25).
Evaluation of the effect of resistance on outcomes and costs is difficult, because the risks of acquiring resistant infection—such as prior antimicrobial therapy, prolonged hospital stay, and admission to intensive care—are themselves associated with poor prognoses. Nevertheless, when adjusted for other risks, mortality rates and length of hospital stay are generally at least twice as great for patients infected with resistant bacteria as it is for those infected with susceptible strains of the same species (26,27,28,29).
In serious infections due to Enterobacteriaceae, resistance to third-generation cephalosporins (which is nearly always associated with multidrug resistance) tends to worsen clinical outcome (30, 31, 32and33). A meta-analysis of bacteremia caused by ESBL-producing Enterobacteriaceae (34) showed significantly increased crude mortality and significantly increased incidence of delay in effective therapy. Mortality rates were higher for infections with imipenem-resistant (K. pneumoniae carbapenemase [KPC]-producing) Enterobacter spp. (11/33) than with susceptible strains (3/33) (35) and 14-day mortality was 9/19 patients with bacteremia due to KPC-producing K. pneumoniae (36). However, a lack of controlled studies limits the assessment of causality in all these studies.
TABLE 86-2 Pooled Means of the Distribution of Antimicrobial Resistance Rates (%) by All ICUs Combined, Non-ICU Inpatient Units and by Outpatients, January 1998 through June 2004
Antibiotic Resistant Pathogen
ICU
Non-ICU
Outpatients
MRSA
52.9
46.0
31.1
Methicillin-resistant CNS
76.6
65.7
50.2
Vancomycin-resistant Enterococcus spp.
13.9
12.0
4.6
Ciprofloxacin-resistant P. aeruginosa
34.8
27.7
23.4
Imipenem-resistant P. aeruginosa
19.1
12.3
7.0
Ceftazidime-resistant P. aeruginosa
13.9
8.8
4.6
Cef3-resistant Enterobacter spp.
27.7
21.0
9.6
Carbapenem-resistant Enterobacter spp.
0.7
1.0
0.5
Cef3-resistant Klebsiella pneumoniae
6.2
5.8
1.8
Cef3-resistant Escherichia coli
1.3
1.5
0.6
Quinolone-resistant E. coli
7.3
8.2
3.6
Penicillin-resistant pneumococci
18.9
18.2
16.8
Ceftriaxone-resistant pneumococci
7.5
7.6
4.8
MRSA, methicillin-resistant S. aureus; Cef3, third-generation cephalosporin; CNS, coagulase-negative staphylococcus.
(From National Nosocomial Infections Surveillance. NNIS report, data summary from January 1992 through June 2004, issued October 2004. Am J Infect Control 2004;32:470-485.)
The impact of drug resistance on the outcome of P. aeruginosa infection has been investigated in a few studies, but the results are not consistent. This may be in part because it is often difficult to distinguish P. aeruginosa colonization from infection and also because invasive disease and bacteremia tend to occur in highly compromised patients with underlying poor prognosis (37, 38, 39and40).
Individual studies of S. aureus infections have shown variable results, but meta-analyses show that infections with MRSA have worse outcomes than those due to methicillin-sensitive S. aureus (MSSA) strains. For example, MRSA bacteremias have twice the mortality of those caused by MSSA (41), and MRSA surgical site infections have significantly greater 90-day mortality, length of hospitalization, and hospital charges (42).
Serious enterococcal infection, especially bacteremia, is associated with severe underlying disease, which itself has a poor prognosis. Some studies find no independent effect of vancomycin resistance on outcomes of enterococcal infections (43,44); others show a significantly worse outcome (measured as treatment failure, attributable mortality, prolonged hospital stay, recurrent bacteremia, endocarditis, ICU admission, surgical intervention, and increased healthcare costs) (45, 46, 47, 48and49).
TABLE 86-3 Mean Resistance Rates in Selected Pathogens Associated with Healthcare-Associated Infections in ICU Patients, January-May 1999 Compared with the 5 Years 1994-1998 and January-December 2003 Compared with the 5 Years 1998-2002
% Increase in Resistance
Antimicrobial/Pathogen
1999 vs. 1994-1998
2003 vs. 1998-2002
Vancomycin/enterococci
47%
12%
Methicillin/Staphylococcus aureus
43%
11%
Methicillin/coagulase-negative staphylococci
2%
1%
3rd Cephalosporin/E. coli
23%
0%
3rd Cephalosporin/K. pneumoniae
-1%
47%
Imipenem/P. aeruginosa
35%
15%
Quinolone/P. aeruginosa
49%
9%
3rd Cephalosporin/P. aeruginosa
<1%
20%
3rd Cephalosporin/Enterobacter spp.
3%
-6%
3rd, third generation.
(From National Nosocomial Infections Surveillance. NNIS report, data summary from January 1990-May 1999, issued June 1999. Am J Infect Control 1999;27:520-532; National Nosocomial Infections Surveillance. NNIS report, data summary from January 1992 through June 2004, issued October 2004. Am J Infect Control 2004;32:470-485.)
The general effect of antimicrobial resistance has been assessed by considering outcomes with “appropriate” or “inappropriate” therapy. Therapy is inappropriate when it is ineffective in vitro against the infecting microorganism or not given promptly by a suitable route (50). Failure to provide prompt, effective antimicrobial therapy may result in treatment failure and death. There is, for example, a significant association between inappropriate treatment and clinical outcome in bacteremia and ventilator-associated pneumonia (51, 52, 53and54).
Delayed response alone is associated with increased morbidity, prolonged hospital stay, additional investigations, procedures and treatments, and increased complications, which, in conditions such as meningitis or bone and joint infection, may result in severe disability. This is costly. Additional costs and lost bed days are incurred by the need to control the spread of antimicrobial-resistant microorganisms within hospitals. Thus, even apart from mortality and morbidity, antimicrobial resistance is hugely expensive in terms of its socioeconomic impact on patients and on the cost-effectiveness of healthcare delivery.
MULTIRESISTANT PROBLEM MICROORGANISMS
Gram-Negative Bacteria
Escherichia coliE. coli is the commonest cause of hospital-acquired gram-negative urinary tract infection (UTI) and septicemia. E. coli is relatively fastidious in its nutritional requirements and does not survive well in the environment and, until recently, did not tend to cause cross-infection or person-to-person spread. For these reasons, most healthcare-associated E. coli infections are endogenous, arising from commensal bowel flora of the affected patient. The species is naturally susceptible to ampicillin but now about 50% to 60% of both healthcare-associated and community isolates are resistant, usually by the production of β-lactamases, enzymes that bind and destroy β-lactam antibiotics. The most common type of β-lactamase in E. coli is TEM-1, accounting for about 80% of such resistance (55, 56and57). TEM-1 is encoded on transferable plasmids and has disseminated throughout the world since its discovery in 1965 (58). Some strains of Enterobacteriaceae, including E. coli, produce TEM-2—a similar enzyme that differs from TEM-1 only in a single amino acid and confers similar phenotypic resistance. Although ampicillin is now unreliable for the treatment of E. coli infection, other drugs usually remain effective, including cephalosporins, quinolones, and aminoglycosides. E. coli can also be treated by the combination of a β-lactam with a β-lactamase inhibitor, such as amoxicillin/clavulanic acid (co-amoxiclav) and ampicillin/sulbactam. The β-lactamase inhibitors prevent the action of TEM-1 or TEM-2 and restore the activity of the β-lactams. This combination may be ineffective, because some E. coli strains can produce excessive amounts of TEM-1 that swamp the effect of the β-lactamase inhibitor (59,60) or are resistant to it (57).
Mutations in TEM-1 and TEM-2 have resulted in new ESBLs that can break down newer cephalosporins, and thus, render E. coli resistant to them. These ESBLs are named TEM-3, TEM-4, etc., and >100 of them have been reported (3,61,). They are often plasmid-borne and are associated with aminoglycoside and other resistances encoded on the same plasmid. However, until recently, E. coli has remained generally antibiotic-sensitive and relatively easy to treat and has not caused much cross-infection or spread within hospitals or the community.
This situation changed with the emergence of E. coli-producing CTX-M ESBLs in the 1990s. CTX-M-producing Enterobacteriaceae first appeared in South America but have now become distributed worldwide (62,63). Compared with other ESBLs, CTX-M enzymes are more active against cefotaxime than against other third-generation cephalosporins. They appear to have originated from the Kluyvera spp. of environmental bacteria, and more than 50 different types have been identified. They are associated with plasmids and transposons and have disseminated widely among the Enterobacteriaceae.
In addition to resistance to penicillins and cephalosporins, CTX-M-producing E. coli are usually also resistant to other previously active agents such as the aminoglycosides and quinolones. Furthermore, these new MDR E. coli appear to be highly transmissible both in the community and in hospitals and have disseminated worldwide. Most isolates are clonally unrelated, but large single strain community outbreaks occur (64). Increasing asymptomatic fecal carriage raises the possibility of spread by food sources and international travel.
This world pandemic of CTX-M-producing E. coli has resulted in a new epidemiology for MDR coliforms (19,65). Opportunistic healthcare-associated outbreaks with mainly single clones of SHV- and TEM-type ESBL-producing K. pneumoniae have been replaced by sporadic and epidemic community infections with heterogenous clones of more virulent MDR CTX-M-producing E. coli. Spread occurs among healthy elderly people at home and in long-term care facilities; admission of these groups to hospital or care homes may result in healthcare-associated outbreaks with consequent further dissemination. Infections tend to occur in the elderly, although younger healthy patients may also be involved. The common presentation is UTI (sometimes complicated by bacteremia) in catheterized, elderly, community, or newly admitted hospital patients. The features of the new community MDR E. coli infections compared with the older MDR KES healthcare-associated infections have been reviewed by Pitout and Laupland (19) and are summarized in Table 86-4.
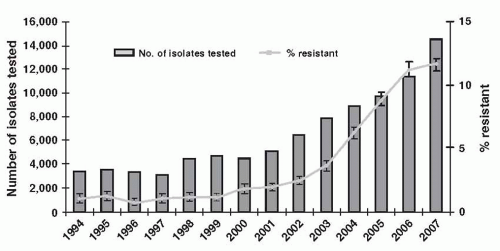
The U.K. Health Protection Agency reviewed E. coli bacteremias reported from hospitals in England, Wales, and Northern Ireland in 2008 (66). There were about 22,000 such bacteremias in 2007. Resistance to third-generation cephalosporins in E. coli blood isolates increased from about 2% in 2001 to 12% in 2007 (Fig. 86-1); resistance to ciprofloxacin rose from 1% to 23%; and resistance to gentamicin rose from 1% to 8.5%. Resistance to carbapenems was rare and was ≤0.2% in 2007. There were far fewer Klebsiella bacteremias—about 6,000 in 2007—and among these, resistance to third-generation cephalosporins was about 14% (up from 4% in 1994); to ciprofloxacin, about 15%; and to gentamicin, about 10%. (Resistance to carbapenems was not reported.) Thus, in 2007, multidrug resistance in UK hospital blood isolates of E. coli was similar to or higher than in Klebsiella, and there were many more of them compared with Klebsiella.
Klebsiella, Enterobacter, and Serratia spp.Klebsiella, Enterobacter, and Serratia spp. are common opportunistic gram-negative pathogens that have similar epidemiologies and clinical presentations. They are all inherently resistant to ampicillin, and Enterobacter spp. and Serratia spp. are resistant to first-generation cephalosporins. These Enterobacteriaceae have a great facility for acquiring and disseminating resistance plasmids and Enterobacter spp. may develop chromosomally mediated resistance to second-and third-generation cephalosporins (67,68).
Enterobacter spp. possess an inducible chromosomally encoded class I β-lactamase that is normally suppressed by a repressor gene and is produced in large amounts only after exposure to certain β-lactams. Full resistance to second- and third-generation cephalosporins results when stably derepressed mutants appear that express the class I β-lactamase constitutively. These mutants are selected by cephalosporin therapy and produce the β-lactamase continuously.
TABLE 86-4 Characteristics of Infections Caused by Extended-Spectrum β-Lactamase-Producing Bacteria
Older Klebsiella spp. etc.
Newer Escherichia Coli
Virulence/place
Less virulent/HAI
More virulent/Community and HAI
Type of ESBL
SHV and TEM types
CTX-M (especially CTX-M15)
Infection
UTI, respiratory, intra-abdominal, blood
Usually UTIs, but also blood, gastroenteritis
Resistances
All β-lactams; usually quinolones and cotrimoxazole; usually aminoglycosides
All β-lactams; usually quinolones and cotrimoxazole; often aminoglycosides
Molecular epidemiology
Most often clonally related
Usually not clonally related, but clonal outbreaks described worldwide, including UK
Risk factors
Longer hospital stay; severity of illness; time in ICU; ventilation; urinary or vascular. Catheterization; previous exposure to antibiotics (especially cephalosporins)
Repeat UTIs/underlying renal pathology; previous antibiotics including cephalosporins and quinolones; previous hospitalization; nursing home residents; older people; diabetes; liver pathology
(From Pitout JD, Laupland KB. Extended-spectrum β-lactamase- producing Enterobacteriaceae: an emerging public-health concern. Lancet Infect Dis 2008;8:159-166.)
To a greater or lesser extent, all three of these species colonize the human bowel and patient’s skin and may spread from person to person via staff members’ hands. They may then go on to colonize the urinary and respiratory tracts of patients treated with β-lactams and may produce bacteremia in the compromised host. They are relatively free-living and can also survive and multiply in nutritionally poor wet environments at room temperature. Because of this, they may contaminate food, enteral feeds, and infusion fluids, leading to widespread common-source outbreaks.
K. pneumoniae is the most frequently isolated member of this group and the most virulent. It is naturally resistant to ampicillin, usually by the production of SHV-1, a β-lactamase similar to TEM-1 and TEM-2, which may be encoded on either the chromosome or, less commonly, on a transferable plasmid (69). The carriage rate for healthy people is low but increases in hospitalized patients, especially during prolonged hospitalization or antibiotic therapy.
During the 1970s, there were frequent reports of hospital outbreaks of gentamicin-resistant K. pneumoniae, sometimes associated with significant mortality when highly compromised patients were involved (70,71). The microorganisms often spread between hospitals and into the community. They became endemic in some hospitals and were sometimes associated with the simultaneous appearance of multiple resistances in other strains of Klebsiella and in other species of Enterobacteriaceae (72,73). In these cases, K. pneumoniae appeared to be acting as an engine of resistance dissemination, especially resistance to aminoglycosides (74).
FIGURE 86-1 Resistance to ceftazidime in E. coli from bacteremias in England, Wales, and Northern Ireland, 1994-2007. (From Health Protection Agency. Antimicrobial resistance and prescribing in England, Wales and Northern Ireland, 2008. London, UK: Health Protection Agency, 2008.)
Once the epidemiology of resistant Klebsiella infection was understood, and following the introduction of newer cephalosporins, these outbreaks became much less common. However, strains of K. pneumoniae (and also Klebsiella ozaenae) then appeared that are resistant to third-generation cephalosporins by the production of ESBLs and can spread to produce hospital outbreaks (61,75, 76, 77and78). The ESBLs are usually the result of mutations in the genes encoding TEM-1, TEM-2, or SHV-1. They are encoded on plasmids that can transfer to other species, and they are often associated with other multiple resistances, including resistance to aminoglycosides (79). Although ESBL-producing strains are usually susceptible to β-lactam-β-lactamaseinhibitor combinations such as amoxicillin/clavulanate and ampicillin/sulbactam, healthcare-associated isolates may be resistant by hyperproduction of the ESBL (76,80). These multiresistant strains may also acquire resistance to quinolones by mutation. Thus, recent isolates are often resistant to all the common β-lactams, aminoglycosides, and quinolones and are reliably susceptible only to the carbapenems. Outbreaks with these new multiresistant Klebsiella spp. have an epidemiology similar to that of the gentamicin-resistant Klebsiella outbreaks of the 1970s.
MDR Klebsiella spp. are now widespread, and some strains have also picked up the CTX-M type ESBLs that have recently emerged in E. coli. For all these Enterobacteriaceae, the treatment of last resort are the carbapenems. The recent emergence of carbapenem resistance is therefore a matter of concern.
Carbapenem resistance in Enterobacteriaceae can be mediated by a variety of mechanisms, including permeability/efflux changes, hyperproduction of ampC β-lactamases or ESBLs, or production of specific carbapenemases. A range of different carbapenemases have been reported, most of them rare or uncommon. They include Ambler class B metallo-β-lactamases such as verona imipenemase and imipenemase; class D OXA-48 β-lactamase, mostly in K. pneumoniae from Turkey, Lebanon, and Belgium; class A clavulanic-acid-inhibited carbapenemases; chromosomally encoded non-metallo-β-lactamases of Serratia spp. and plasmid-mediated KPCs (81).
In Europe, carbapenem-resistant Enterobacteriaceae are rare (except in Greece), but KPC-producing bacteria are now widespread in certain parts of China, Israel, Greece, South America, and the United States (36,82).
Only gold members can continue reading. Log In or Register to continue