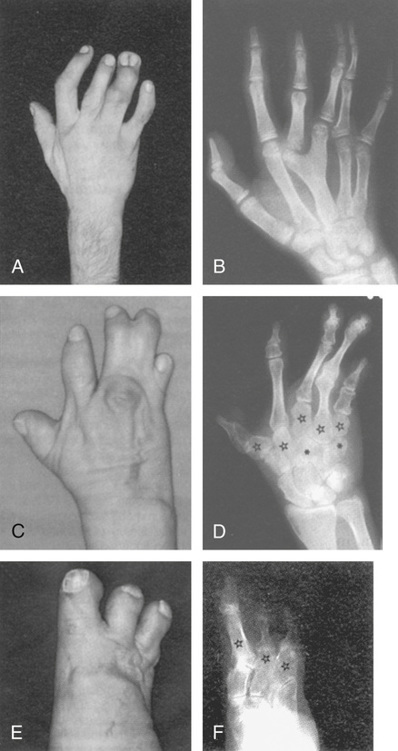
The importance of transcription factors in normal development is illustrated by an unusual mutation of HOXD13 that causes synpolydactyly, an incompletely dominant condition in which heterozygotes have interphalangeal webbing and extra digits in their hands and feet. Rare homozygotes have similar but more severe abnormalities and also have bone malformations of the hands, wrists, feet, and ankles (Fig. 14-17). The HOXD13 mutation responsible for synpolydactyly is caused by expansion of a polyalanine tract in the amino-terminal domain of the protein; the normal protein contains 15 alanines, whereas the mutant protein contains 22 to 24 alanines. The polyalanine expansion that causes synpolydactyly is likely to act by a gain-of-function mechanism (see Chapter 11), as heterozygosity for a HOXD13 loss-of-function mutation has only a mild effect on limb development, characterized by a rudimentary extra digit between the first and second metatarsals and between the fourth and fifth metatarsals of the feet. Regardless of the exact mechanism, this condition demonstrates that a general function for HOX genes is to determine regional identity along specific body axes during development.
Morphogens and Cell to Cell Signaling
One of the hallmarks of developmental processes is that cells must communicate with each other to develop proper spatial arrangements of tissues and cellular subtypes. This communication occurs through cell signaling mechanisms. These cell-cell communication systems are commonly composed of a cell surface receptor and the molecule, called a ligand, that binds to it. On ligand binding, receptors transmit their signals through intracellular signaling pathways. One of the common ligand-receptor pairs is the fibroblast growth factors and their receptors. There are 23 recognized members of the fibroblast growth factor gene family in the human, and many of them are important in development. The fibroblast growth factors serve as ligands for tyrosine kinase receptors. Abnormalities in fibroblast growth factor receptors cause diseases such as achondroplasia (Case 2) (see Chapter 7) and certain syndromes that involve abnormalities of craniofacial development, referred to as craniosynostoses because they demonstrate premature fusion of cranial sutures in the skull.
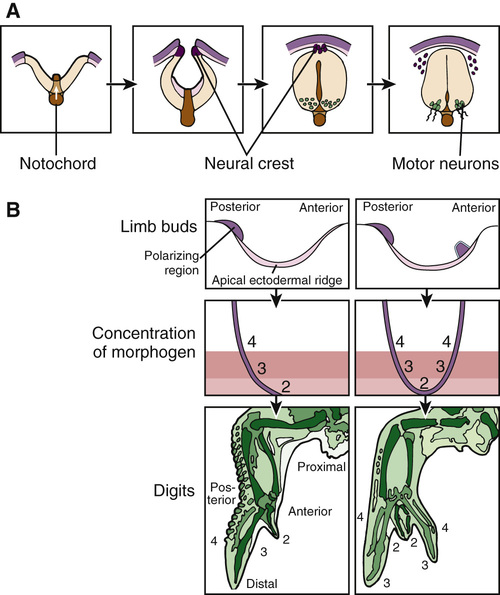
One of the best examples of a developmental morphogen is hedgehog, originally discovered in Drosophila and named for its ability to alter the orientation of epidermal bristles. Diffusion of the hedgehog protein creates a gradient in which different concentrations of the protein cause surrounding cells to assume different fates. In humans, several genes closely related to Drosophila hedgehog also encode developmental morphogens; one example is the gene sonic hedgehog (SHH). Although the specific programs controlled by hedgehog in Drosophila are very different from those controlled by its mammalian counterparts, the underlying themes and molecular mechanisms are similar. For example, secretion of the SHH protein by the notochord and the floor plate of the developing neural tube generates a gradient that induces and organizes the different types of cells and tissues in the developing brain and spinal cord (Fig. 14-18A). SHH is also produced by a small group of cells in the limb bud to create what is known as the zone of polarizing activity, which is responsible for establishing the posterior side of the developing limb bud and the asymmetrical pattern of digits within individual limbs (see Fig. 14-18B).
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


