Anterolateral abdominal muscles
The three muscle layers of the body wall (see p. 181) are separate in the flanks, where they are known as the external oblique, internal oblique and transversus abdominis muscles. The layers have fused ventrally to form the rectus abdominis muscle.
External oblique
The muscle arises by eight digitations, one from each of the lower eight ribs just lateral to their anterior extremities. The lower four slips interdigitate with the costal fibres of latissimus dorsi and the upper four with digitations of serratus anterior. From its fleshy origin the muscle fans out to a very wide insertion, much of which is aponeurotic. The muscle has a free posterior border which extends from the twelfth rib to its insertion by fleshy fibres into the anterior half of the outer lip of the iliac crest. Muscular fibres are replaced by an aponeurosis below a line joining the anterior superior iliac spine to the umbilicus, and medial to a vertical line drawn from the tip of the ninth costal cartilage (Fig. 5.2). The limit of the fleshy fibres is visible in an athlete as a graceful curve. The aponeurotic fibres, directed obliquely downwards and forwards, interdigitate with each other across the front of the rectus abdominis along the whole length of the linea alba. (This description is adequate for all practical purposes although detailed studies of cadaveric material have revealed that the aponeurotic fibres are in superficial and deep layers, the fibres in the superficial layer running obliquely upwards and those in the deep layer at right angles downwards. The fibres continue across the midline after decussation, the fibres from the deep layer passing to the superficial layer on the contralateral side of the abdominal wall and vice versa.) The free horizontal upper border of this aponeurosis extends from the fifth rib to the xiphisternum. It is the only structure in the anterior sheath of the rectus muscle above the costal margin.
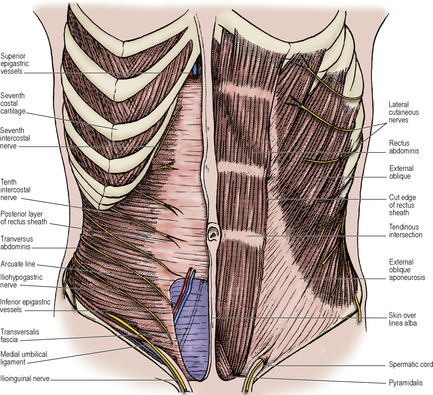
The free posterior border of the muscle forms the anterior boundary of the lumbar triangle (of Petit) that is floored in by the internal oblique and bounded behind by the anterior border of latissimus dorsi and below by the iliac crest. The triangle may be the site of a rare lumbar hernia (see Fig. 2.4, p. 40).
The lower border, lying between the anterior superior iliac spine and the pubic tubercle, forms the inguinal ligament (of Poupart). Its edge is rolled inwards to form a gutter; the lateral part of this gutter gives origin to part of the internal oblique and transversus abdominis muscles. The fascia lata of the thigh is attached to the inguinal ligament and when the thigh is extended the fascia lata pulls the inguinal ligament downwards into a gentle convexity.
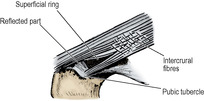
Just above and lateral to the pubic tubercle is an oblique, triangular gap, the superficial inguinal ring, in the aponeurosis (Fig. 5.3). The base of the gap is the pubic crest, and the margins are the crura of the ring.
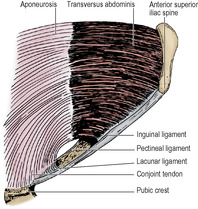
From the medial end of the inguinal ligament the triangular lacunar ligament (of Gimbernat) extends horizontally backwards to the pectineal line on the pubis (see Fig. 3.1, p. 112). A fibrous band, the pectineal ligament (of Astley Cooper) extends laterally further along the pectineal line. The crescentic free lateral edge of the lacunar ligament is the medial margin of the femoral ring (see p. 118).
From the pubic tubercle, fibres that may be traced upwards and medially, behind the spermatic cord, interdigitate in the linea alba with those of the opposite side. This is the reflected part of the ligament (Fig. 5.3). Near the apex of the superficial inguinal ring are fibres running at right angles to those of the aponeurosis, the intercrural fibres, that prevent the crura from separating.
Internal oblique
Fleshy fibres of the muscle arise from the whole length of the lumbar fascia, from the intermediate area of the anterior two-thirds of the iliac crest and from the lateral two-thirds of the inguinal ligament. From the lumbar fascia the muscle fibres run upwards along the costal margin, to which they are attached, becoming aponeurotic at the tip of the ninth costal cartilage. Below the costal margin, the aponeurosis splits around the rectus muscle, the two layers rejoining at the linea alba. Halfway between the umbilicus and the pubic symphysis the posterior layer ends in a curved free margin, the arcuate line. Below this point, the aponeurosis passes wholly in front of the rectus muscle, to the linea alba (Fig. 5.6) (but see p. 224).
The muscle fibres that arise from the inguinal ligament are continued into an aponeurosis that is attached to the crest of the pubic bone and, more laterally, to the pectineal line (Fig. 5.4). This aponeurosis is fused with a similar arrangement of the transversus aponeurosis to form the conjoint tendon. The internal oblique therefore has a free lower border, which arches over the spermatic cord: laterally the margin consists of muscle fibres in front of the cord; medially the margin consists of tendinous fibres behind the cord.
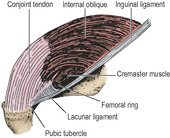 |
| Figure 5.4 Left conjoint tendon and lacunar ligament. The lowest fibres of the internal oblique arise from the inguinal ligament and arch medially to reach the conjoint tendon, forming as they do so the roof of the inguinal canal. They cover up the similar fibres of transversus abdominis, shown in Figure 5.5. Note that the nearly vertical conjoint tendon lies at right angles to the nearly horizontal lacunar ligament. Only the upper part of the cremaster muscle is depicted. |
Transversus abdominis
The muscle arises in continuity from the lateral third of the inguinal ligament, the anterior two-thirds of the inner lip of the iliac crest, the lumbar fascia, the twelfth rib, and from the inner aspects of the lower six costal cartilages where it interdigitates with the diaphragm. The muscle fibres become aponeurotic and pass behind the rectus to fuse with the internal oblique aponeurosis in the linea alba. Below the arcuate line the aponeurosis passes wholly in front of the rectus muscle. (As in the case of external oblique, detailed cadaveric studies have shown that the aponeurotic fibres of transverse abdominis that contribute to the rectus sheath are in two layers at right angles to each other.) In the upper part of the abdomen the outer margin of the aponeurosis is more medial, and muscular fibres lie behind the lateral part of rectus abdominis. The lower fibres of the aponeurosis curve downwards and medially with those of the internal oblique as the conjoint tendon, to insert on the pubic crest and the pectineal line (Fig. 5.5).
Rectus abdominis and pyramidalis
Rectus abdominis arises by two heads: a medial from in front of the pubic symphysis and a lateral from the upper border of the pubic crest. The lower parts of the two muscles are narrower and lie edge to edge. The upper parts are broader and are separated from each other by the linea alba (Fig. 5.2). They are inserted on to the front of the fifth to seventh costal cartilages. Typically three tendinous intersections are found in the muscle, one at the umbilicus, one at the xiphisternum, and one between these two; one or two incomplete intersections are sometimes found below the umbilicus. The tendinous intersections blend inseparably with the anterior layer of the rectus sheath. They occupy only the superficial part of the rectus and do not penetrate to the posterior surface of the muscle, which is thus not connected to the posterior layer of the sheath. The contracting rectus abdominis can be seen as bulgings between the tendinous intersections in an individual who is not too fat.
The small triangular pyramidalis muscle arises from the body of the pubis and the symphysis between rectus abdominis and its sheath. It converges with its fellow into the linea alba 4cm or so above its origin.
Between the two recti all the aponeuroses that form the rectus sheath fuse to form the linea alba, a strong midline fibrous structure which is firmly attached to the xiphoid process above and the pubic symphysis below (Fig. 5.2). Above the symphysis it is very narrow, for here the two recti are in contact with one another behind it. From just below the umbilicus to the xiphisternum it broadens out between the recti. Here the fibres form a tough felted membrane. The umbilicus is a defect in the linea alba through which fetal umbilical vessels pass.
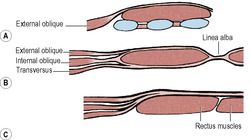
Rectus sheath
The aponeurosis of the internal oblique splits into anterior and posterior layers to enclose the rectus muscle (Fig. 5.6B). The external oblique aponeurosis fuses with the anterior layer to form the anterior layer of the sheath, and the transversus aponeurosis fuses with the posterior layer to form the posterior layer of the sheath. From halfway between the umbilicus and the pubic symphysis all three aponeuroses pass in front of the muscle (Fig. 5.6C). The aponeuroses of internal oblique and transversus fuse completely but that of the external oblique fuses only to the most medial part of the sheath. The posterior layer of the sheath has a free lower margin concave downwards, the arcuate line or semicircular line (of Douglas). Superiorly the posterior layer of the sheath is attached to the costal margin (seventh, eighth and ninth costal cartilages). Above the costal margin the anterior layer of the sheath consists only of the external oblique aponeurosis (Fig. 5.6A).
The splitting of the internal oblique aponeurosis along the lateral border of the rectus muscle forms a relatively shallow groove, the semilunar line. It curves up from the pubic tubercle to the costal margin at the tip of the ninth costal cartilage in the transpyloric plane.
Detailed studies indicate that the aponeuroses of external oblique, internal oblique and transversus abdominis are each bilaminar, giving six layers in all; three form the anterior and three the posterior layers of the rectus sheath. These layers decussate across the midline. There may not be a well-defined arcuate line but a gradual diminution of aponeurotic fibres with increasing thickness of the transversalis fascia. The lower thickened part of the transversalis fascia, between the iliac crest and pubis just above the inguinal ligament, is called the iliopubic tract.
Contents. Apart from the rectus and pyramidalis muscles, the sheath contains the ends of the lower six thoracic nerves and their accompanying posterior intercostal vessels, and the superior and inferior epigastric vessels.
The intercostal nerves (T7–11; see p. 183) pass from their intercostal spaces into the abdominal wall between the internal oblique and transversus muscles, and run round in this neurovascular plane to enter the sheath by piercing the posterior layer of the internal oblique aponeurosis. They then proceed behind the rectus muscle to about its midline (Fig. 5.2), where they pierce the muscle, supply it, and pass through the anterior layer of the sheath to become the anterior cutaneous nerves. In the sheath T7 runs upwards just below the costal margin, T8 transversely and the others obliquely downwards. Before they reach the sheath the nerves give off their lateral cutaneous branches, which pierce the internal and external oblique to reach the skin.
The lowest thoracic nerve, T12 or subcostal, is described on page 278.
The superior epigastric artery, a terminal branch of the internal thoracic (see p. 184), enters the sheath by passing between the sternal and highest costal fibres of the diaphragm. It supplies the rectus muscle and anastomoses within it with the inferior epigastric artery. This vessel leaves the external iliac at the inguinal ligament (Fig. 5.8), passes upwards behind the conjoint tendon, slips over the arcuate line and so enters the sheath. Veins accompany these arteries, draining to internal thoracic and external iliac veins respectively.
A pedicled flap of the upper part of the rectus muscle based on the superior epigastric artery—or a free flap of the lower part with anastomosis of the divided inferior epigastric artery to the internal thoracic artery—is used in reconstructive breast surgery.
Blood supplies
Apart from the intercostal and epigastric vessels mentioned above, the anterolateral abdominal muscles also receive a blood supply from the lumbar and deep circumflex iliac arteries. The lumbar arteries are described on page 276; they end among the flat anterolateral muscles and do not reach the rectus sheath.
The deep circumflex iliac artery arises from the external iliac (see p. 276) behind the inguinal ligament (Fig. 5.8), and runs laterally towards the anterior superior iliac spine in a sheath formed by the transversalis and iliac fasciae where they meet. It continues along the inner lip of the iliac crest, pierces the transversus muscle to reach the neurovascular plane and anastomose with branches of the iliolumbar and superior gluteal arteries. At the anterior superior iliac spine it gives off an ascending branch which may be at risk in a gridiron incision (see p. 232).
Lymph drainage
The superficial tissues of the anterolateral abdominal wall drain in quadrants: to the pectoral group of axillary nodes above the umbilicus on each side, and to superficial inguinal nodes below that level. The deeper parts of the wall drain into vessels in the extraperitoneal tissues. Above the umbilicus these pierce the diaphragm to reach mediastinal nodes, and below it they run to the external iliac and para-aortic nodes.
Nerve supplies
The rectus muscle and external oblique are both supplied by the lower intercostal and subcostal nerves (T7–T12), and the internal oblique and transversus by those same nerves but with the addition of the iliohypogastric and ilioinguinal nerves (L1). The lowest fibres of the internal oblique and transversus that continue medially as the conjoint tendon receive the L1 innervation, which thus helps to maintain the integrity of the inguinal canal (see below). Pyramidalis is supplied by the subcostal nerve (T12).
Actions of abdominal muscles
The muscles of the anterior abdominal wall have four main roles: (1) to move the trunk, (2) to depress the ribs (expiration), (3) to compress the abdomen (evacuation, expiration, heavy lifting), and (4) to support the viscera (intestines only). The abdominal wall, moving to and fro with breathing, conforms to the volume of the abdominal contents. Its shape is determined by the tonus of its own muscles. The subumbilical pull of healthy flank muscles keeps its lower part flat by holding back the lower recti.
Moving the trunk. As the muscles are attached to the thoracic cage and the bony pelvis their action is to approximate the two. They are flexors of the vertebral column in its lumbar and lower thoracic parts. Rectus abdominis is the most powerful flexor. The oblique muscles are also lateral flexors and rotators of the trunk.
Depressing the ribs. The recti and obliques approximate the ribs to the pelvic girdle. If erector spinae prevents thoracolumbar flexion this provides a powerful expiratory force (e.g. coughing, blowing the trumpet). Added to this is the abdominal compression (aided by transversus) that elevates the diaphragm to increase the expiratory effort.
Compressing the abdomen. While flexion of the vertebral column is prevented by the erector spinae muscles, the oblique muscles compress the abdominal cavity; in this they are aided strongly by transversus abdominis, which has no flexing action on the spine. The recti play little part in compression. If the diaphragm is relaxed, it is forced up, as in expiration. At the same time levator ani helps to hold the pelvic effluents closed. The reverse occurs in evacuation of the pelvic effluents. Here the diaphragm contracts to resist upward displacement, but it is a far weaker muscle than the abdominal wall, and in forceful compression it is prevented from rising by holding the breath, i.e. by closure of the glottis, and perhaps of the mouth and nostrils (see p. 395).
Supporting and protecting viscera. If the anterior abdominal wall is incised or removed, only the intestines spill out. The upper abdominal viscera, such as the liver, spleen and kidneys, do not require the support of the wall. Reflex contraction in response to a blow helps to protect all viscera.
Tests. Rectus abdominis can be tested by lying flat on the back and raising the head (without using the arms). There are no specific tests for the other flat muscles. The abdominal reflex and Beevor’s sign have been referred to on page 17.
Inguinal canal
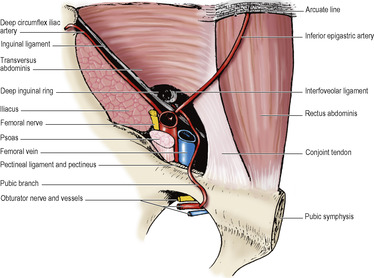
The inguinal canal is an oblique intermuscular slit about 4cm long lying above the medial half of the inguinal ligament. It commences at the deep inguinal ring, ends at the superficial inguinal ring, and transmits the spermatic cord and ilioinguinal nerve in the male and the round ligament of the uterus and ilioinguinal nerve in the female. Its anterior wall is formed by the external oblique aponeurosis (Fig. 5.2), assisted laterally by the internal oblique muscle (Fig. 5.7). Its floor is the inrolled lower edge of the inguinal ligament, reinforced medially by the lacunar ligament (Fig. 5.4). Its roof is formed by the lower edges of the internal oblique and transversus muscles, which arch over from in front of the cord laterally to behind the cord medially, where their conjoined aponeuroses, constituting the conjoint tendon, are inserted into the pubic crest and the pectineal line of the pubic bone. The posterior wall of the canal is formed by the strong conjoint tendon medially and the weak transversalis fascia throughout.
The integrity of the inguinal canal depends upon the strength of the anterior wall in the lateral part and of the posterior wall in the medial part, provided the abdominal muscles are of good tone and their aponeuroses unyielding. The deep and superficial inguinal rings lie at opposite ends of the inguinal canal and the intervening part of the canal is pressed flat when the aponeuroses are under tension and the intra-abdominal pressure raised. The conjoint tendon lies posterior to the superficial inguinal ring and helps to reinforce this area. Laterally the transversalis fascia in the posterior wall is strengthened by the presence in front of it of tendinous, and sometimes muscular, fibres derived from the transversus abdominis muscle. These fibres constitute the interfoveolar ligament (Fig. 5.8). They arch down from the lower border of transversus around the vas to the inguinal ligament, and constitute the functional medial edge of the deep ring.
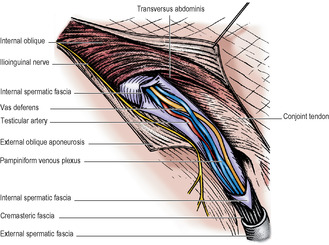
The deep inguinal ring lies about 1.25cm above the midpoint of the inguinal ligament and is an opening in the transversalis fascia. From the margins of this opening the transversalis fascia is projected along the canal, like a sleeve, the internal spermatic fascia, around the structures that pass through the ring. These are the vas deferens and its artery, the testicular artery and the accompanying veins (usually double at this level, Fig. 5.10), the obliterated remains of the processus vaginalis, the genital branch of the genitofemoral nerve, autonomic nerves and lymphatics. These structures constitute the spermatic cord; in the female they are replaced by the obliterated processus vaginalis, the round ligament and lymphatics from the uterus. The ilioinguinal nerve, although a content of the inguinal canal, does not enter the canal through the deep ring, but by piercing the internal oblique muscle, i.e. it slips into the canal from the side, not from the back. The nerve lies in front of the cord and leaves the canal through the superficial ring to supply skin of the inguinal region, upper part of the thigh, anterior third of the scrotum (or labium majus) and root of the penis.
 |
| Figure 5.10 |
Structures deep to the posterior wall
Crossing the posterior wall at the medial edge of the deep inguinal ring is the inferior epigastric artery. Lateral to the artery the vas deferens in the male and the round ligament of the uterus in the female enter the canal by hooking around the interfoveolar ligament. At the deep ring the inferior epigastric artery gives off the cremasteric branch to supply that muscle and the coverings of the cord. The area bounded laterally by the inferior epigastric artery, medially by the lateral border of the rectus muscle, and below by the inguinal ligament is the inguinal triangle (of Hesselbach). By definition a hernial sac passing lateral to the artery (i.e. through the deep ring) is an indirect hernia, one passing medial to the artery (through the inguinal triangle) is a direct hernia; the latter stretches out the conjoint tendon over itself and is therefore seldom large. As an inguinal hernia emerges through the superficial inguinal ring it lies above and medial to the pubic tubercle, while the neck of a femoral hernia (see p. 118) is below and lateral to the pubic tubercle.
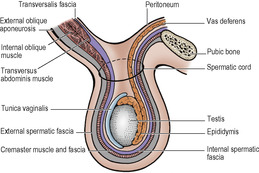
Spermatic cord
The spermatic cord has three coverings and six (groups of) constituents.
Of the three coverings of the spermatic cord (Fig. 5.9), the internal spermatic fascia is derived from the transversalis fascia at the deep inguinal ring. As the cord passes through the ring into the inguinal canal, it picks up a second covering, the cremaster muscle and cremasteric fascia. This loosely arranged layer consists of striated muscle bundles united by areolar tissue. The muscle arises laterally from the inguinal ligament, the internal oblique and transversus abdominis muscles. The fibres spiral down the cord (the longest reaching as far as the tunica vaginalis of the testis) and loop back to become attached to the pubic tubercle. The third covering, the external spermatic fascia, is acquired from the external oblique aponeurosis as the cord passes between the crura of the superficial ring.
The cremaster muscle can elevate the testis towards or even into the inguinal canal; although the fibres are skeletal the action is reflex rather than voluntary. This cremasteric reflex is particularly active in the infant and child and must be borne in mind when examining the scrotum in the young, to avoid an erroneous diagnosis of undescended testis.
The constituents of the cord consist of:
• The vas deferens, which usually lies in the lower and posterior part of the cord.
• Arteries, the largest of which is the testicular artery (see below), with the artery to the vas (from the superior or inferior vesical), and the cremasteric artery (from the inferior epigastric, Fig. 5.8) to the coverings.
• Veins—the pampiniform plexus (see below).
• Lymphatics, essentially those from the testis draining to para-aortic nodes, but including some from the coverings which drain to external iliac nodes.
• Nerves, in particular the genital branch of the genitofemoral nerve which supplies the cremaster muscle. Other nerves are sympathetic twigs which accompany the arteries.
• The processus vaginalis, the obliterated remains of the peritoneal connection with the tunica vaginalis of the testis. When patent it forms the sac of an indirect inguinal hernia.
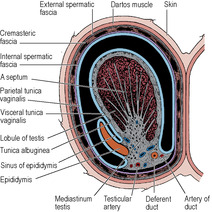
Testis
The testis (Fig. 5.12) is an oval organ possessing a thick covering of fibrous tissue, the tunica albuginea. The epididymis is attached to its posterolateral surface; this is an important point to remember when trying to distinguish between swellings of these two structures. The vas deferens arises from the lower pole of the epididymis (see p. 231) and runs up medial to it behind the testis. The front and sides of the testis lie free in a serous space formed by the overlying tunica vaginalis, a remnant of the fetal processus vaginalis. This serous membrane covers also the anterolateral part of the epididymis and lines a slit-like space, the sinus of the epididymis, which lies between testis and epididymis. Testis, epididymis and tunica vaginalis lie in the scrotum surrounded by thin membranes, adherent to each other, that are downward prolongations of the coverings of the spermatic cord (Fig. 5.9). Right and left sides are separated by the median scrotal septum (see p. 321). Average testicular dimensions are 5cm (length), 2.5cm (breadth), 3cm (anteroposterior diameter). The appendix testis is a minute sessile cyst attached to the upper pole of the testis within the tunica vaginalis. It is a remnant of the paramesonephric duct (see p. 304).
Blood supply
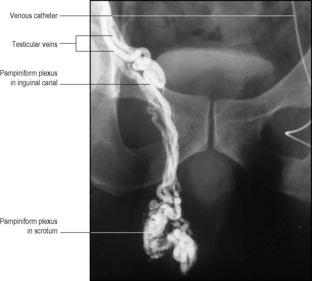
The testicular artery, from the aorta, runs in the spermatic cord, gives off a branch to the epididymis, and reaches the back of the testis, where it divides into medial and lateral branches. These do not penetrate the mediastinum testis (see below), but sweep around horizontally within the tunica albuginea. Branches from these vessels penetrate the substance of the organ. In the region of the epididymis there is an anastomosis between the testicular, cremasteric and ductal arteries; but if the main artery is divided, the smaller vessels may not completely sustain the testis and atrophy may occur, though ischaemic necrosis is unlikely. Venules reach the mediastinum, from which several veins pass upwards in the spermatic cord as a mass of intercommunicating veins, the pampiniform plexus (Fig. 5.10), which surround the testicular artery. In the inguinal canal the plexus separates out into about four veins which join to form two that leave the deep inguinal ring, becoming single on psoas major on the posterior abdominal wall. The left vein invariably joins the left renal vein at a right angle and the right drains directly into the inferior vena cava at an acute angle. The testicular veins usually have valves. Varicocele (varicosities of the pampiniform and cremasteric veins) occurs much more frequently on the left side than the right.
Lymph drainage
Lymphatics from the testis run back with the testicular artery to para-aortic nodes lying alongside the aorta at the level of origin of the testicular arteries (L2 vertebra), i.e. just above the umbilicus. The testicular lymph therefore does not drain to inguinal nodes, although the overlying scrotal skin does.
Nerve supply
The testis is supplied by sympathetic nerves. Most of the connector cells lie in T10 segment of the cord. Passing mainly in the lesser splanchnic nerve to the coeliac ganglia the efferent fibres synapse there. Postganglionic grey fibres reach the testis along the testicular artery. Sensory fibres share the same sympathetic pathway. They run up along the testicular artery and through the coeliac plexus and lesser splanchnic nerve and its white ramus to cell bodies in the posterior root ganglion of T10 spinal nerve.
Structure
The upper pole of the epididymis is attached high up on the posterolateral surface of the testis. Here there is a fibrous mass, the mediastinum testis, from which septa radiate to reach the tunica albuginea. The septa divide the testis into some 200–300 lobules, each of which contains 1–4 highly convoluted seminiferous tubules. The cut surface of the organ bulges with protruding tubules. The seminiferous tubules open into the rete testis, which is a network of intercommunicating channels lying in the mediastinum testis. From the rete 12–20 vasa efferentia enter the commencement of the canal of the epididymis, thus attaching the head of the epididymis to the testis.
The seminiferous tubules have several layers of cells. The outermost layer consists of spermatogonia, which divide to produce the primary spermatocytes. These divide to form secondary spermatocytes. They have a very short life and divide almost immediately to form spermatids. These do not divide but undergo a metamorphosis into spermatozoa. The whole process of producing spermatozoa from spermatogonia is termed spermatogenesis.
Among the developing germ cells are the supporting or sustentacular cells (of Sertoli). The Sertoli cells secrete an androgen binding protein (ABP) which keeps a high concentration of testosterone in the germ cell environment.
Scattered among the cells of the connective tissue between the tubules (outside them) are the interstitial cells (of Leydig). Larger than fibroblasts, they constitute the endocrine portion of the testis and secrete testosterone.
Development and descent of the testis
The testis develops from the gonadal ridge, formed by proliferation of the coelomic epithelium and a condensation of underlying mesoderm, on the medial side of the mesonephros (see p. 23). Primordial germ cells from the yolk sac migrate to the gonadal ridge and become incorporated in the developing gonad. At first the testis and mesonephros are situated on the posterior abdominal wall, attached by the urogenital mesentery. As the testis enlarges its cranial end degenerates and the remaining organ lies at a more caudal location. Most of the mesonephros atrophies. Derivatives of the remaining mesonephric tubules include the vasa efferentia of the testis and the paradidymis (a small collection of tubules above the epididymis at the lower end of the spermatic cord). In the male, the mesonephric duct forms the canal of the epididymis, vas deferens, ejaculatory duct and the appendix of the epididymis (a small appendage on the head of the epididymis).
A condensation of mesodermal cells, the gubernaculum, connects the lower pole of the testis to the region of the anterior abdominal wall that later forms the scrotum (Fig. 5.11). It traverses the site of the future inguinal canal, which is formed around it by the developing muscles of the abdominal wall. A sac of peritoneum, the processus vaginalis, protrudes down the inguinal canal anterosuperior to the gubernaculum. By the seventh month of fetal life the testis is in the deep inguinal ring and thereafter it progresses rapidly through the inguinal canal into the scrotum before birth. As the testis descends it is accompanied by the processus vaginalis. The testis projects into the distal part of the processus, which forms the tunica vaginalis. The rest of this peritoneal sac usually gets obliterated. Persistence of the whole, or proximal part, of the sac maintaining its connection with the peritoneal cavity constitutes a hernial sac, a clinical hernia occurring when intra-abdominal contents enter the sac. Persistence of an intervening segment of the processus may lead to the development of a hydrocele of the cord. Accumulation of serous fluid between the layers of the tunica vaginalis forms the much more common hydrocele of the testis.
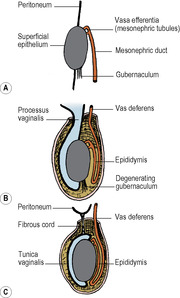
Sometimes the testis is not fully descended at birth, but enters the scrotum during the first few months thereafter. Failure to descend may result in cryptorchid testis, where it remains in the abdomen, or descent may be arrested anywhere from the deep inguinal ring downwards. Undescended testes are peculiarly liable to malignant disease; spermatogenesis is defective or absent but androgenic activity is not. They must be distinguished from retracted testes, where the cremaster muscle draws them back into the canal, especially in the young under the influence of cold examining hands!
Epididymis and vas deferens
The epididymis is a firm structure, attached behind the testis, with the vas deferens to its medial side. It consists of a single highly coiled tube packed together by fibrous tissue. It has a large head at its upper end, connected by a body to a pointed tail at its lower end. The head is connected to the upper pole of the testis by the vasa efferentia and the tail to the lower pole by loose connective tissue. The body is partly separated from the testis by a recess which is open laterally, the sinus of the epididymis (Fig. 5.12). The lateral surface of the epididymis is covered by the tunica vaginalis, which also lines the sinus.
From the tail the vas deferens, a direct continuation of the canal of the epididymis, provided with a thick wall of smooth muscle, passes up medially. It enters the spermatic cord, passes through the inguinal canal, across the side wall of the pelvis just under the peritoneum, and crosses the pelvic cavity. It pierces the prostate and opens by the ejaculatory duct into the prostatic urethra. Its pelvic course is described on page 300.
Blood supply
The epididymis is supplied by a branch of the testicular artery. This enters the upper pole and runs down to the lower pole. It anastomoses with the tiny artery to the ductus.
Venous and lymphatic drainage are as for the testis.
Nerve supply
The epididymis is supplied, like the testis, by sympathetic fibres from the coeliac ganglion via the testicular artery.
Structure
The epithelial lining of the coiled tube that forms the epididymis is columnar in type, with long microvilli called stereocilia. The thin wall has a single layer of circular smooth muscle.
Development
The whole length of the single tube constituting the epididymis and vas is a persistent and much elongated part of the mesonephric (Wolffian) duct of the embryo. This duct receives the efferent tubules of the mesonephros (see p. 285). When the mesonephros is replaced by the metanephros and disappears, some of its tubules persist and attach to the developing testis, forming the vasa efferentia and draining the products of the testis into the commencement of the mesonephric duct. Some mesonephric tubules persist without serving any function of drainage. Thus, above and below the epididymis blind tubules, the vasa aberrantia, open into its canal. Their bulbous blind ends may form small swellings; an upper one is relatively constant, the appendix of the epididymis. Above the epididymis, at the lower extremity of the spermatic cord, a mass of tubules, blind at each end, persists as the paradidymis (organ of Giraldès). A cyst formed from an aberrant tubule will contain spermatozoa and thus be opalescent. A cyst formed from a tubule of the paradidymis cannot contain spermatozoa, and its fluid is thus crystal clear.
The paramesonephric (Müllerian) duct, developing into the uterine tube and uterus (see p. 304), disappears in the male except at its two ends. The upper end persists as the appendix testis, the conjoined lower ends of the two ducts persist as the prostatic utricle (utriculus masculinus) (see p. 299).
Vasectomy
The spermatic cord containing the firm tubular vas is palpated between the thumb and fingers at the top of the scrotum and a transverse incision made so that the vas can be dissected out and a small length of it removed. Each remaining cut end is turned back on itself and ligated, and the same procedure is then carried out on the opposite side.
Abdominal incisions
The simplest abdominal incision is the midline incision, above or below the umbilicus (or both, skirting the umbilicus) and passing through skin and subcutaneous tissues, the linea alba, transversalis fascia, extraperitoneal fat and peritoneum. No major vessels or nerves are involved, but a few small vessels may cross the midline of the peritoneum. In the lower abdomen the linea alba is very narrow and the two rectus muscles lie very close together; here poor suture technique predisposes to incisional hernias. In the suprapubic region the bladder must not be damaged.
For laparoscopic surgery, the incision for insertion of a needle to induce pneumoperitoneum was usually made in the midline, just above or below the umbilicus, and the instrument was first directed down towards the pelvic cavity to avoid damaging the aorta. Increasingly now the pneumoperitoneum is created by trocat and cannula insertion through an umbilical port, which is also used for camera insertion. Other ports for instruments lateral to the rectus sheath must not be made too low, to avoid the inferior epigastric vessels (see Fig. 4.1, p. 180); transillumination from within the peritoneal cavity helps to avoid them. The development of multifunctional instruments has enabled some minimally invasive procedures (such as appendicectomy and cholecystectomy) to be performed entirely via an umbilical port. Laparoscopic extraperitoneal inguinal and femoral hernia repair utilises pre-peritoneal balloon inflation to create the required surgical space. Sites below the umbilicus or at the lateral border of the rectus sheath are used for the insertion of a trocar and cannula for the drainage of peritoneal fluid (paracentesis) or a peritoneal dialysis catheter. A suprapubic catheter to drain a distended bladder is usually introduced through the midline.
In a paramedian incision, the anterior wall of the rectus sheath is incised vertically 2cm from the midline and the rectus muscle retracted laterally so that the posterior wall of the sheath can be incised. The tendinous intersections in the rectus muscle at and above the umbilicus have to be dissected off the anterior wall of the sheath; they may contain vessels. Above the umbilicus on the right the falciform ligament may have to be divided. In a rectus split incision, through a vertical incision 3cm from the midline the rectus is split instead of being retracted. The small part of the muscle medial to the split will be denervated and devascularized but this usually does not cause problems. The lack of a posterior wall of the sheath below a point midway between the umbilicus and pubic symphysis implies that sound healing depends on proper closure of the sheath’s anterior wall.
The right subcostal (Kocher’s) incision is made 3cm parallel to and below the right costal margin, from the midline to beyond the lateral border of the rectus sheath. The incision is often made more horizontal than parallel to the subcostal margin. The anterior layer of the sheath (with the external oblique) and the rectus muscle are divided in the line of the skin incision, with ligation of the superior epigastric vessels and/or their branches. The posterior layer of the sheath is then incised, continuing laterally into the internal oblique and transversus and through to the peritoneum. The seventh intercostal nerve follows the costal margin upwards and is usually above the incision line, but the eighth or ninth nerve may have to be cut, with little effect on the rectus muscle. Cutting more than two nerves (paralysing more of the rectus) should be avoided.
The double Kocher or curved rooftop incision, combining subcostal incisions on both sides, gives a very wide exposure of the upper abdomen.
The gridiron (McBurney’s) incision is a right lower oblique muscle-splitting incision, long used for appendicectomy. The skin incision runs downwards and medially through the junction of the outer and middle thirds of a line drawn from the anterior superior iliac spine to the umbilicus. The external oblique muscle and its aponeurosis are divided in the line of their fibres and then the internal oblique and transversus are split transversely (in the line of their own fibres). The two muscles are close and may be split together. The transversus becomes aponeurotic at this level and some fibres may pass to the transversalis fascia. The peritoneum can then be incised. The iliohypogastric and ilioinguinal nerves may be seen between the internal oblique and transversus and must not be damaged, to avoid weakening the protective effect that the muscles exert upon the inguinal canal. Extending the incision laterally may cut the deep circumflex iliac artery’s ascending branch, which runs upwards above the anterior superior iliac spine between the internal oblique and transversus.
For cosmetic reasons the gridiron incision is often replaced by a more transverse muscle-splitting incision in a skin crease starting above and medial to the anterior superior iliac spine and extending nearly to the lateral border of the rectus sheath.
The oblique muscle-cutting incision (Rutherford Morison’s) is similar to the gridiron but after incising the external oblique in the line of its fibres the internal oblique and transversus are cut in the same line (not in the line of their own fibres).
Transverse muscle-cutting incisions can be made at or about the level of the umbilicus, cutting the rectus sheaths and the obliques and transversus muscles. The rectus muscle is retracted medially or divided. Lower intercostal nerves run obliquely through the abdominal wall, but more than one is not likely to be cut by this incision.
The lower abdominal transverse incision (Pfannenstiel’s) is commonly used for approach to the pelvic organs. A skin crease incision is made above the pubic symphysis, just below the hairline, as far as the lateral borders of the rectus sheaths. The anterior layers of the rectus sheaths are divided in the line of the skin incision and flaps dissected off the muscles both upwards and downwards, with the pyramidalis muscles included in the lower flap. The rectus muscles which at this level lie close together are separated to expose transversalis fascia which is incised with the peritoneum, care being taken to avoid the bladder. Transverse division of the rectus muscles gives wider exposure, and the incisions can be extended laterally into the flat muscles.
A lumbar incision is used for extraperitoneal approach to the kidney and upper ureter. The incision extends below the twelfth rib from the lateral border of erector spinae towards the anterior superior iliac spine. Latissimus dorsi and external oblique are incised and their cut edges retracted so that the internal oblique and transversus merging with the lumbar fascia can also be incised. The subcostal nerve deep to internal oblique should be preserved but the vessels can be ligated. The transversalis fascia and extraperitoneal fat in the posterior part of the incision are separated to expose the renal fascia. The peritoneal cavity is not entered. Proper identification of the twelfth rib is essential to avoid entering the pleural cavity, which extends below its medial part (see p. 213).
Part two. Abdominal cavity
The abdominal cavity is much more extensive than the impression gained from examination of the anterior abdominal wall. Much of it lies under cover of the lower ribs, for the domes of the diaphragm arch high above the costal margin. Hidden by the lower ribs are the liver and spleen, much of the stomach, and the upper poles of the kidneys and both suprarenals. The volume of the thoracic cavity is, correspondingly, much less than examination of the bony thorax would suggest. Furthermore, an appreciable amount of the abdominal cavity projects backwards into the pelvis, just in front of the buttocks. A perforating wound of the buttock can easily involve the pelvic cavity. The pelvic cavity accommodates not only its own pelvic organs (rectum, uterus, bladder, etc.), but also a goodly volume of intestine (sigmoid colon and ileum).
General topography of the abdomen
The alimentary canal and its two chief derivatives the liver and pancreas (and also the spleen) are developed in fetal mesenteries which later alter their disposition as a result of fusion of adjacent leaves of peritoneum. The liver and spleen remain invested in peritoneum, but the pancreas becomes retroperitoneal.
The alimentary canal is invested unevenly. Parts of it are suspended in the abdominal cavity by peritoneal folds (‘mesenteries’); other parts become plastered down to the posterior abdominal wall. The stomach is fixed at its two ends, but is elsewhere suspended by ‘mesenteries’. The duodenum is plastered down to the posterior abdom-inal wall, while the whole length of the small intestine swings free on its own mesentery. The ascending and descending colon are both adherent to the posterior abdominal wall, but between the colic flexures the transverse colon is mobile on its own mesentery, the transverse mesocolon. The sigmoid (pelvic) colon swings free on a mesentery, while the rectum is plastered by peritoneum to the hollow of the sacrum.
The suprarenals, kidneys and ureters lie behind the peritoneum. The aorta and inferior vena cava also lie behind the peritoneum, and intestinal vessels run through the mesenteries to reach the gut.
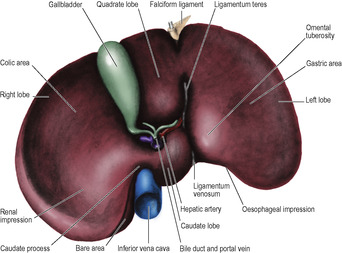
The transpyloric plane bisects the body between the jugular notch and the pubic symphysis. This level is approximately midway between the xiphisternum and the umbilicus, or about a hand’s breadth below the xiphisternal joint (Fig. 5.1). It cuts each costal margin at the tip of the ninth costal cartilage, which is at the lateral border of the rectus abdominis (semilunar line); deep to this point on the right side lies the fundus of the gallbladder. The plane passes through the lower border of the first lumbar vertebra, where the spinal cord ends at the conus medullaris.
As its name implies, the plane usually passes through the pylorus, but the pylorus is suspended by the lesser and greater omenta, and is therefore relatively mobile. The plane passes along the head, neck and body of the pancreas, just above the attachment of the transverse mesocolon. The supracolic compartment (liver, spleen, fundus of stomach) lies above the plane, the infracolic compartment (small intestine, colon) below it. The superior mesenteric artery leaves the aorta, and the splenic vein joins the superior mesenteric vein to form the portal vein at this level. The hilum of each kidney lies at the plane, the right just below and the left just above it.
Part three. Peritoneum
The peritoneum is a serous membrane which lines the abdominal cavity; it covers the anterior and posterior walls, the undersurface of the diaphragm and the walls and floor of the pelvic cavity. All this is the parietal peritoneum. In places it leaves the posterior abdominal wall or diaphragm or pelvic floor to form a partial or complete investment for viscera; this is the visceral peritoneum, which forms the serous covering for many viscera.
Peritoneum consists of a single layer of flattened cells, with phagocytic properties, overlying areolar tissue which varies in both thickness and density in different places. Over expansile parts this areolar tissue is loose and cellular (e.g. transversalis fascia on the anterior abdominal wall) while over non-expansile parts it is often thick (e.g. iliac fascia, psoas fascia, parietal pelvic fascia); but loose or dense, thin or thick, these variously named fasciae are part of the one continuous extraperitoneal connective tissue lying between the parietal peritoneum and the walls of the abdominal and pelvic cavities.
Various folds or reflexions of peritoneum connect viscera to the abdominal walls or to one another. Some of these are properly called folds, but others are called mesentery, omentum or ligament. The double fold supporting the small intestine is the mesentery; the mesenteries supporting the transverse colon, sigmoid colon and appendix are the transverse mesocolon, sigmoid mesocolon and mesoappendix. The lesser omentum connects the stomach to the liver, and the greater omentum hangs down from the lower border of the stomach. The various ligaments associated with the liver, stomach and spleen are simply peritoneal folds attached to them, and the broad ligament stretches out on either side of the uterus.
Peritoneal folds of the anterior abdominal wall
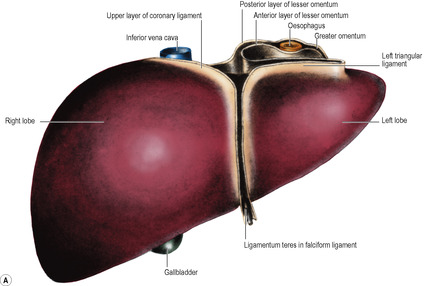
On the posterior surface of the anterior abdominal wall the peritoneum is raised into six folds, one above and five below the umbilicus. The falciform ligament consists of two adherent layers of peritoneum connecting the anterior surface of the liver to the supraumbilical part of the anterior abdominal wall, just to the right of the midline, and to the inferior surface of the diaphragm. Its concave, inferior margin, which contains the ligamentum teres (the obliterated remains of the left umbilical vein, see p. 31), deviates to the right and is attached to the notch for this ligament on the inferior border of the liver.
Below the umbilicus there is a central fold with a pair on either side. Centrally is the median umbilical fold, containing the median umbilical ligament (the obliterated remains of the urachus; see p. 298). On each side, and also running as far as the umbilicus, is the medial umbilical fold, containing the medial umbilical ligament (the obliterated remains of the umbilical artery; see p. 31). Farther laterally is the lateral umbilical fold, containing the inferior epigastric vessels, which enter the rectus sheath by passing across the arcuate line; although called umbilical folds, this lateral pair do not reach as far as the umbilicus.
Peritoneal cavity: greater and lesser sacs
The serous-coated organs fill the abdominal cavity so that visceral surfaces are in contact with one another or with the parietal peritoneum. The space between them is only potential, not actual, and it contains only a few millilitres of tissue fluid which lubricates adjacent surfaces so they can glide over one another. This is the general peritoneal cavity or greater sac.
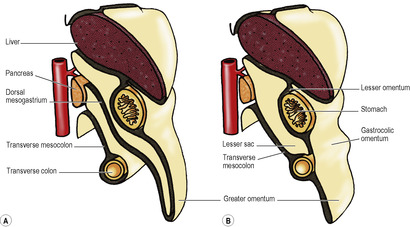
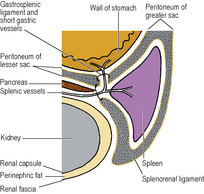
The omental bursa, or lesser sac, is a subsection or diverticulum of the peritoneal cavity behind the stomach. It opens into the greater sac through a slit-like aperture in front of the inferior vena cava, the epiploic foramen (see p. 236). The anterior wall of the lesser sac is formed by the posterior layer of the lesser omentum, the peritoneum over the posterior aspect of the stomach and the posterior of the anterior two layers of the greater omentum (Fig. 5.13). The posterior wall is formed by the anterior of the two posterior layers of the greater omentum which adheres to, but is surgically separable from, the anterior surface of the transverse colon and the transverse mesocolon. Above the attachment of the transverse mesocolon to the anterior border of the pancreas, the posterior wall is formed by the peritoneum that covers the front of the neck and body of pancreas, upper part of left kidney, left suprarenal gland, commencement of abdominal aorta, coeliac artery (plexus and nodes) and part of the diaphragm (Fig. 5.14). Theoretically the cavity of the lesser sac should extend down between the anterior two layers and the posterior two layers of the greater omentum, but because of fusion of these layers the cavity does not extend much below the transverse colon. The narrow upper border of the lesser sac is at the right side of the abdominal oesophagus, where the peritoneum of the posterior wall is reflected anteriorly on the inferior aspect of the diaphragm to form the posterior layer of the lesser omentum. At the tail end of the pancreas the left border of the lesser sac is formed by the splenorenal and gastrosplenic ligaments (see below and Fig. 5.49).
Greater omentum
The greater omentum is a double sheet of peritoneum, folded on itself to form four layers (Fig. 5.13). The anterior two layers descend from the greater curvature of the stomach (where they are continuous with the peritoneum on the anterior and posterior surfaces of the stomach) like an apron, overlying coils of intestine, and then turn round and ascend up to the transverse colon where they loosely blend with the peritoneum on the anterior surfaces of the transverse colon and the transverse mesocolon above it. The four layers of the greater omentum below the transverse colon fuse with each other to form an integral structure. This contains adipose tissue of variable amount, depending on the nutritional status of the patient and numerous macrophages.
The part of the greater omentum between the stomach and the transverse colon is often referred to as the gastrocolic omentum. The right and left gastroepiploic vessels run between the layers of the gastrocolic omentum, close to the greater curvature of the stomach. The lesser sac may be accessed through the gastrocolic omentum. Other routes of surgical access to the lesser sac are through the lesser omentum and through the transverse mesocolon.
Below the stomach the left border of the greater omentum envelops the spleen, except for a small bare area at the hilum. The spleen therefore lies in the general peritoneal cavity. Two double-layered folds of peritoneum, the gastrosplenic and splenorenal ligaments, connect the hilum of the spleen to the greater curvature of the stomach and the anterior surface of the left kidney respectively. The splenic vessels and pancreatic tail lie in the splenorenal ligament and the short gastric and left gastroepiploic vessels run in the gastrosplenic ligament (Fig. 5.49).
Lesser omentum
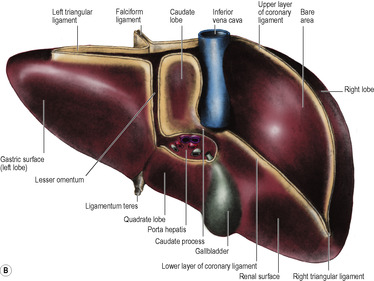
The two layers of peritoneum that extend between the liver and the upper border (lesser curvature) of the stomach constitute the lesser omentum or gastrohepatic omentum). It can usually only be seen when the liver is lifted up, away from the stomach. Its attachment to the stomach extends from the right side of the abdominal oesophagus and along the lesser curvature to the first 2cm of the duodenum (Fig. 5.35). The liver attachment is L-shaped (Fig. 5.32B), to the fissure for the ligamentum venosum and the porta hepatis. Between the duodenum and the liver it has a right free margin, where the anterior and posterior layers of peritoneum become continuous. This fold forms the anterior boundary of the epiploic foramen.
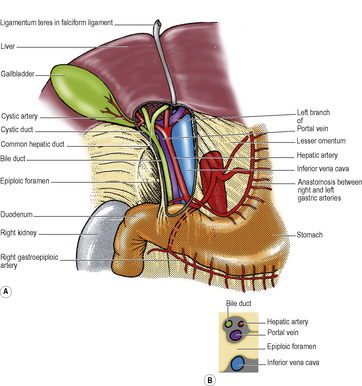
The epiploic foramen (of Winslow, or the aditus to the lesser sac, Fig. 5.35) is a vertical slit about 2.5cm at the right border of the lesser sac. Its upper boundary is the caudate process of the liver (Fig. 5.32B). The lower boundary is the first part of the duodenum. The posterior boundary is the inferior vena cava, covered by the parietal peritoneum of the posterior abdominal wall which, continuing to the left through the foramen, becomes the peritoneum of the posterior wall of the lesser sac. Anteriorly the foramen is bounded by the right free margin of the lesser omentum containing between its two peritoneal layers the portal vein, and anterior to it the hepatic artery and bile duct, with the duct to the right of the artery, as well as autonomic nerves, lymphatics and nodes.
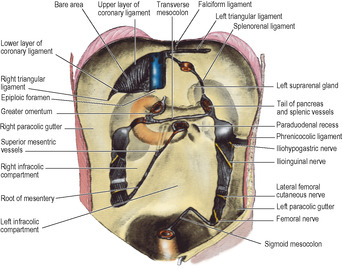
Peritoneal compartments
The peritoneal cavity is descriptively divided into compartments called supracolic, infracolic and pelvic.
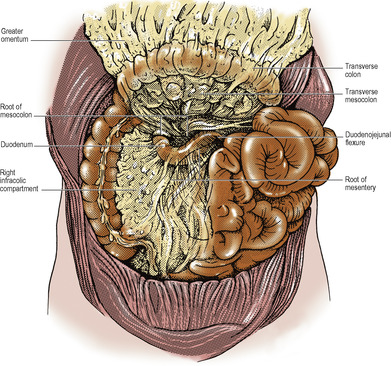
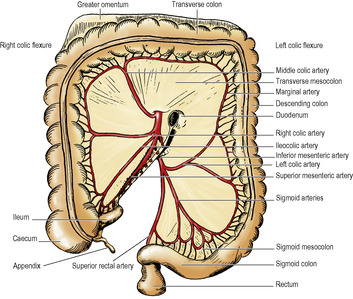
The dividing line between the supracolic and infracolic compartments is the attachment of the transverse mesocolon to the posterior abdominal wall, or rather to the organs that lie on the abdominal wall at this level (Figs 5.13 and 5.14). The transverse mesocolon is a double fold of peritoneum passing from the transverse colon to the front of the second part of the duodenum, and to the anterior aspect of the head and the anterior border of the body of the pancreas. The transverse colon and transverse mesocolon are adherent to the posterior surface of the greater omentum. When the greater omentum is lifted up over the costal margin, the stomach, transverse colon and mesocolon are lifted upwards with it, and the posterior surface of the mesocolon and the infracolic compartment brought into view (Fig. 5.15).
The attachments of the liver to the diaphragm and abdominal wall define the subdivisions of the supracolic compartment (Figs 5.16 and 5.32). To the right and left of the falciform ligament are the right and left subphrenic (subdiaphragmatic) spaces. These two spaces are closed above by the superior layer of the coronary ligament and the anterior layer of the left triangular ligament respectively. Behind the right lobe of the liver and in front of the right kidney is the right subhepatic space or hepatorenal pouch (of Morison). This space is closed above by the inferior layer of the coronary ligament and the small right triangular ligament. To the right it is bounded by the abdominal surface of the diaphragm. On the left side the space communicates through the epiploic foramen with the lesser sac or left subhepatic space. Below it is continuous with the right paracolic gutter (see below).
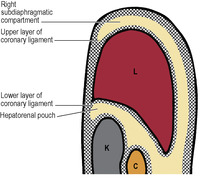
When lying supine, the hepatorenal pouch is the lowest part of the peritoneal cavity (with the sole exception of the pelvis), and hence is an area where intraperitoneal fluid of any sort is likely to accumulate.
The infracolic compartment, below the level of the transverse mesocolon, is divided into two by the attachment of the root of the mesentery of the small intestine (Fig. 5.14), which passes down from left to right at an angle of about 45°. It begins on the left at the duodenojejunal junction, crosses the third part of the duodenum where the superior mesenteric vessels enter between its two layers, and then continues downwards across the aorta, inferior vena cava, right psoas muscle and ureter to the right iliac fossa. This attachment is 15cm long. The intestinal border of the mesentery is plicated like the hem of a very full skirt and measures about 6m long. The depth of the mesentery (from root to gut) is greatest at the central part, about 20cm.
In the retroperitoneal tissue in the region of the root of the mesentery there are numerous Pacinian corpuscles. Tension and traction on peritoneal folds in the upper abdomen produce a fall of blood pressure by undue stimulation of these encapsulated mechanoreceptors.
To the right of the root of the mesentery is the triangular right infracolic space (Fig. 5.14). Its apex lies below, at the ileocaecal junction. Its right side is the ascending colon, and its upper border is the attachment of the transverse mesocolon.
Lateral to the ascending colon is the right paracolic gutter. It can be traced upwards into the hepatorenal pouch and downwards into the pelvis—pathways for the gravitation of fluid.
The left infracolic space is larger than the right infracolic compartment and is quadrilateral in shape. It widens below where it is continuous across the pelvic brim with the cavity of the pelvis (Fig. 5.14). Its upper border is the attachment of the transverse mesocolon, and its left side is the descending colon.
Lateral to the descending colon is the left paracolic gutter (Fig. 5.14). It is limited above by a small transverse fold of peritoneum between the left (splenic) flexure of the colon and the diaphragm, the phrenicocolic ligament. Traced downwards the gutter leads to the left of the attachment of the lateral limb of the sigmoid mesocolon at the pelvic brim.
At the lower end of the left infracolic compartment is the attachment of the sigmoid mesocolon (Fig. 5.14). It is Λ-shaped and the two limbs diverge from each other at the bifurcation of the common iliac vessels, on the pelvic brim over the left sacroiliac joint. The lateral limb passes forwards along the pelvic brim (over the external iliac vessels) halfway to the inguinal ligament, while the medial limb slopes down into the hollow of the sacrum, where it reaches the midline in front of S3 vertebra, at the commencement of the rectum. At the apex of the attachment of the pelvic mesocolon, just beneath the peritoneum and lying over the bifurcation of the common iliac artery, is the left ureter, with the inferior mesenteric vessels medial to it, the vein lying between the ureter and the artery.
Nerve supply
The parietal peritoneum is supplied segmentally by the spinal nerves that innervate the adjacent muscles. Thus the diaphragmatic peritoneum is supplied centrally by the phrenic nerve (C4)—hence referred pain and hyperaesthesia from this area to the tip of the shoulder. The remainder of the parietal peritoneum is supplied segmentally by intercostal and lumbar nerves. In the pelvis the obturator nerve is the chief source of supply. The visceral peritoneum is innervated by afferent nerves which travel with the autonomic supply to the viscera. Pain from diseased viscera is due to ischaemia, muscle spasm and stretching of the visceral peritoneum, including mesenteric folds or involvement of the parietal peritoneum.
Retroperitoneal space
Several major structures lie on the posterior abdominal wall behind the peritoneum. These include the aorta and inferior vena cava with a number of their branches and tributaries; the cisterna chyli, lymph nodes and vessels; nerves (mostly branches of the lumbar plexus) including the sympathetic trunks; the kidneys, ureters, pancreas, ascending and descending colon and most of the duodenum and suprarenal glands. All these can be said to lie in the retroperitoneal space, though the term is often used to apply only to the area of the posterior abdominal wall behind the peritoneum that is not occupied by the major viscera and great vessels, e.g. over parts of psoas and other muscles. Haemorrhage and infection may develop in it and blood and pus may be confined to the retroperitoneal space.
Part four. Development of the gut
The disposition of the gut and its mesenteries in the early embryo is simple. The more complex arrangement in the adult is due to elongation and consequent coiling of the alimentary canal and to fusion of certain adjacent peritoneal surfaces.
Gut arteries
Just before the sixth week of embryonic life the alimentary canal is a simple tube passing through to the hind end, its whole length supported by a dorsal mesentery attached in the midline in front of the aorta (Fig. 5.17). Three gut arteries leave the aorta and pass ventrally to supply the tube. The most cranial passes in the dorsal mesogastrium to supply the foregut, the next passes through the dorsal mesentery to supply the midgut and the last passes through the dorsal mesocolon to supply the hindgut. They are the coeliac, the superior mesenteric and the inferior mesenteric arteries respectively, and they continue to supply the derivatives of these parts of the alimentary canal in the adult.
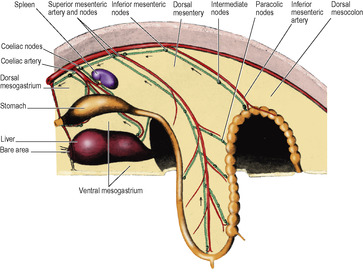
The foregut possesses, in addition, a ventral mesogastrium (derived from the septum transversum, see p. 24) attached in the midline to the undersurface of the diaphragm and the anterior abdominal wall down to the umbilicus (Fig. 5.17). Its caudal free edge is crescentic and carries the left umbilical vein.
The derivatives of the foregut (liver and pancreas) and also the spleen are supplied with blood by the artery of the foregut, the coeliac artery. The liver develops as an outgrowth from the foregut at its junction with the midgut. A tube grows ventrally into the ventral mesogastrium, bifurcates, and cells proliferate from the blind end of the two divisions to form the two lobes of the liver, which are thus enclosed between the two layers of the ventral mesogastrium. The pancreas develops as two outgrowths, one into the ventral and one into the dorsal mesogastrium. These two parts subsequently fuse and exchange ducts by anastomosis, but the double origin of the pancreas is imprinted on the adult by the persistence of two pancreatic ducts, the main and the accessory. The spleen develops by proliferation of cells in the left leaf of the dorsal mesogastrium. It is not a derivative of the foregut itself. The coeliac artery supplies the stomach and these three organs in the adult, where it is properly called the coeliac trunk.
Lymph drainage
Lymph from the alimentary canal drains by lymph vessels that run with the arteries and end ultimately in lymph nodes that lie in front of the aorta at the roots of the three gut arteries. Lymph from the mucous membrane of the alimentary canal passes through several filters. In the mucous membrane itself, from tonsils to anal margin, are lymphoid follicles. In the mesentery, at its gut margin, are lymph nodes (the ‘para-’ group, e.g. paracolic). In the mesentery between its gut margin and root are further intermediate nodes. The preaortic nodes, inferior mesenteric, superior mesenteric, and coeliac, are interconnected by lymph vessels. All the lymph thus ultimately reaches the coeliac nodes, whence it passes to the cisterna chyli (see p. 247). This simple lymphatic arrangement persists in the adult.
Herniation of the gut
By the end of the sixth week the liver has enlarged greatly and the gut has elongated, both to such an extent that the more leisurely growing abdominal walls cannot accommodate them. A loop of gut extrudes into the umbilical cord as the physiological hernia (Fig. 5.17). The loop remains in the umbilical cord for a full month. At the end of the tenth week the abdominal walls have grown enough to accommodate the abdominal contents and the hernia is reduced.
The herniated loop of gut is that supplied by the superior mesenteric artery and it is defined as the midgut. It is destined to produce all the small intestine from the distal part of the duodenum (i.e. distal to the entry of the bile duct) and the proximal part of the colon, almost as far as the left colic flexure. The apex of the loop is at the attachment of the vitellointestinal duct (see p. 24), the site of the ileal (Meckel’s) diverticulum. The main trunk of the superior mesenteric artery is directed to the apex of the loop. Many branches run from it to the proximal limb of the loop, extending from the ventral pancreatic bud to Meckel’s diverticulum. They persist as the jejunal and ileal branches. Only three branches run to the distal limb of the loop; all three persist in the adult as the ileocolic, right colic and middle colic arteries. Their directions are altered considerably after the reduction of the physiological hernia and the rotation of the gut.
Rotation of the midgut
As the loop of midgut in the physiological hernia returns to the abdominal cavity it rotates so that the distal limb goes up on the left and the proximal limb goes down on the right, i.e. to the observer looking at the front of the abdomen, in an anticlockwise direction (Fig. 5.18). The distal loop, developing into colon, thus comes to lie anterior to the commencement of the proximal loop. This part of the proximal loop becomes, after some rotation, plastered to the posterior abdominal wall as the duodenum, and the mesentery of the transverse colon thus comes to lie across it (Fig. 5.19). The last part of the midgut to be reincluded within the abdominal cavity is the caecum, which lies first near the midline, high up. It grows then to the right, turns downwards at the right colic flexure and stops elongating at the right iliac fossa. It leaves a trail of large intestine to indicate its migration and drags the attached lower end of the ileum with it.
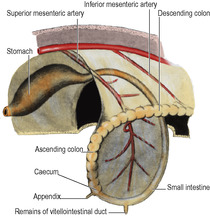
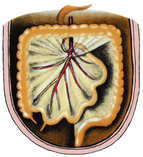
Rotation of the midgut loop occurs around the axis of the superior mesenteric artery, so in the adult the branches to the proximal loop (jejunal and ileal arteries) come off its left side while the three branches to the distal loop (colic arteries) leave its right side (Figs 5.19 and 5.23).
The simple dorsal mesentery of the midgut containing the superior mesenteric artery is, of course, twisted and distorted during the return of the rotated loop of midgut and the subsequent migration of the caecum. Its attachment to the proximal loop causes it to pass across the posterior abdominal wall from the commencement of the loop (duodenum) to the ileocaecal junction (Fig. 5.14). The dorsal mesentery of the distal loop of the midgut hinges like a door across from the midline to the right. It comes into contact with the parietal peritoneum in the right paravertebral gutter, with which it fuses, the colic vessels lying immediately deep to it and in front of everything else on the posterior abdominal wall. The dorsal mesentery of the most distal part of the distal loop, pulled across transversely, does not fuse completely with the parietal peritoneum and persists, with the middle colic artery between its layers, as the transverse mesocolon (Fig. 5.19).
Movement of the hindgut
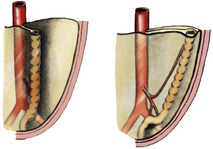
As the midgut loop returns to the abdominal cavity, the hindgut swings on its dorsal mesocolon like a door across to the left (Fig. 5.20), and the mesocolon fuses with the parietal peritoneum of the left paravertebral gutter; a white line (of Toldt) marks the site of fusion along the left side of the descending colon. Hence the left colic vessels lie in front of everything else on the posterior abdominal wall. At the pelvic brim fusion of the layers is not complete and a part of the dorsal mesocolon of the hindgut remains free as the sigmoid mesocolon of the adult.
Growth of the liver
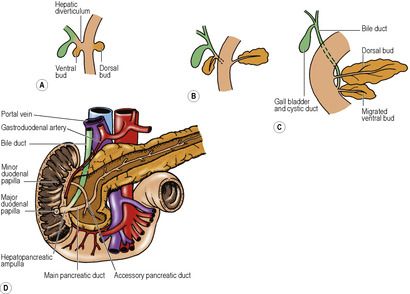
The liver grows apace, and soon outstrips the ventral mesogastrium in which it lies. It grows caudally into the free edge of the ventral mesogastrium, which it pushes down until the left umbilical vein (ligamentum teres in the adult) notches its inferior border and is enclosed in a deep groove on its undersurface. This encroachment of the liver over the crescentic free margin of the ventral mesogastrium divides the latter into two separate parts, namely the falciform ligament between liver and anterior abdominal wall and the lesser omentum (gastrohepatic omentum) between liver and stomach (Fig. 5.17). The lesser omentum is attached to the liver along a fissure that runs backwards behind the fissure for the ligamentum teres (Fig. 5.33), and lodges the fibrous remnant of the ductus venosus (ligamentum venosum).
Rotation of the foregut
Coincident with the growth of the liver, the foregut rotates. The liver originally was ventral to the foregut, in the ventral mesogastrium, and both lie in the midline. As the liver grows it swings to the right, taking the ventral mesogastrium with it. The stomach swings across to the left and in doing so rotates (Fig. 5.21). It has already elongated and broadened, with its dorsal border becoming convex, and its ventral border concave. The distal end of the foregut, destined to become the duodenum (i.e. proximal to the entry of the bile duct), does not dilate in this manner, and its dorsal mesentery shortens. The duodenal part of the gut elongates into a loop which swings to the right and becomes plastered to the posterior abdominal wall (like the ascending and descending parts of the colon). At the same time its walls grow asymmetrically so that the ventral bile duct and pancreatic duct are carried around to open on the medial wall (Fig. 5.40) in line with the duct of the dorsal diverticulum. The duodenum is now fixed in position; so, too, is the oesophagus at the diaphragm. Between these two fixed points as an axis, the dorsal convexity of the stomach rotates to the left. The dorsal convexity becomes the greater curvature, and the original left side of the stomach now faces anteriorly. The concave ventral border becomes the lesser curvature, attached by the lesser omentum (originally the ventral mesogastrium) to the liver, and to the diaphragm between liver and oesophagus. The original right surface of the stomach now lies behind, and faces the peritoneum of the posterior abdominal wall. The space behind the stomach and the lesser omentum is the lesser sac, and the free edge of the lesser omentum lies over the opening from the greater sac into this space. The rotation of the stomach accounts for the left and right vagal nerve trunks becoming anterior and posterior, respectively, in relation to the stomach.
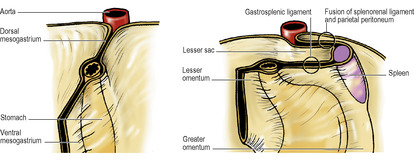
Meanwhile changes have occurred in the disposition of the dorsal mesogastrium. As the dorsal border of the stomach swings to the left the dorsal mesogastrium hinges to the left from this attachment and adheres to the parietal peritoneum as far left as the front of the left kidney. From the front of the left kidney, and from the diaphragm above it, the two layers of the dorsal mesogastrium pass to the oesophagus and the upper part of the greater curvature of the stomach. They form part of the greater omentum, and constitute the left boundary of the lesser sac behind the stomach. The spleen projects from the left leaf into the greater sac, and it divides this part of the greater omentum into gastrosplenic and splenorenal ligaments (Fig. 5.21).
The more caudal part of the dorsal mesogastrium is attached to the lower part of the greater curvature of the stomach, the inferior border of the pylorus and the first 2cm of the duodenum. Its dorsal attachment, in the midline, hinges to the left and becomes plastered to, and fused with, the peritoneum on the posterior abdominal wall over the pancreas. Below this it balloons down like an apron from the greater curvature, over the transverse mesocolon and transverse colon, and returns to the posterior abdominal wall (Fig. 5.13). The deeper part of the double layer fuses with the upper leaf of the transverse mesocolon and with the transverse colon itself, forming the inferior limit of the lesser sac. The superficial part hangs down from the greater curvature to the transverse colon, to which it adheres, forming the gastrocolic omentum, a part of the greater omentum, which is the anterior wall of the lower part of the lesser sac.
From the transverse colon the dependent dorsal mesogastrium hangs down over the front of the coils of small intestine as a fat-containing apron, the main greater omentum.
Part five. Vessels and nerves of the gut
Blood supply of the foregut
Coeliac trunk
This is the artery of the foregut, and its three branches—the left gastric, splenic and common hepatic arteries—supply not only the gut from the lower part of the oesophagus down to the opening of the bile duct into the duodenum, but also the foregut derivatives (the liver and pancreas) and the spleen. It arises from the front of the abdominal aorta between the crura of the diaphragm a little below the median arcuate ligament, at the level of the body of T12 vertebra. It is usually a short wide trunk, flanked by the coeliac group of preaortic lymph nodes. The coeliac ganglia lie one on each side and they send to the artery sympathetic nerves which are carried along all its branches.
At the upper border of the pancreas the trunk divides into its three branches behind the peritoneum of the posterior wall of the lesser sac (Fig. 5.22).
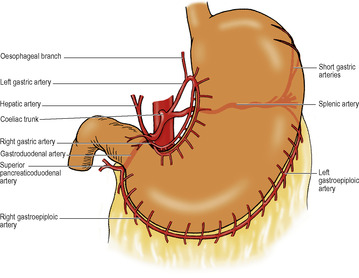
The left gastric artery runs upwards across the left crus towards the oesophageal opening in the diaphragm. It gives off oesphageal branches and turns anteroinferiorly, raising a small fold of peritoneum, the left gastropancreatic fold. It then runs to the right in the lesser omentum along the lesser curvature and supplies the stomach.
The splenic artery passes to the left. It is characteristically very tortuous; the crests of its waves appear above the pancreas, and the troughs lie hidden behind its upper border. It runs across the left crus and left psoas to the hilum of the left kidney, where it turns forward in the splenorenal ligament to the hilum of the spleen. Apart from the spleen it is the main supply to the pancreas. Before breaking up into its terminal splenic branches it gives off about 6 short gastric arteries which run in the gastrosplenic ligament, and the left gastroepiploic artery which runs to the right in the greater omentum, a little distance away from the greater curvature, from where it gives branches to the stomach and the omentum. From the middle part of its course the splenic artery may give off a posterior gastric artery to the stomach, which raises a fold of parietal peritoneum, the gastrosplenic ligament, as it arches forwards towards the stomach.
The common hepatic artery passes over the upper border of the pancreas, downwards and to the right behind the peritoneum of the posterior abdominal wall as far as the first part of the duodenum. It then turns forward, raising a small fold of peritoneum, the right gastropancreatic fold, and curves upwards between the two layers of the lesser omentum as the hepatic artery. Here it meets the bile duct and lies on its left side, both in front of the portal vein surrounded by the peritoneum at the free edge of the lesser omentum. On reaching the porta hepatis, the hepatic artery divides into right and left branches to supply the right and left halves of the liver. These branches and the associated aberrant or accessory hepatic arteries are described on page 262.
The common hepatic usually gives off the right gastric and gastroduodenal arteries.
The right gastric artery leaves the common hepatic as it turns forwards into the lesser omentum. It runs to the left along the lesser curvature and anastomoses with the left gastric artery.
The gastroduodenal artery passes down behind the first part of the duodenum, where it may be eroded by a duodenal ulcer. At the lower border of the duodenum it divides into two. The right gastroepiploic artery passes forward between the first part of the duodenum and the pancreas, and turns to the left between the two leaves of the greater omentum. It runs close to the greater curvature of the stomach and anastomoses with the left gastroepiploic artery.
The other branch of the gastroduodenal artery is the superior pancreaticoduodenal artery. It divides into a smaller anterior and a larger posterior branch, which may arise directly from the gastroduodenal artery; they anastomose with similar branches of the inferior pancreaticoduodenal branch of the superior mesenteric artery. The pancreaticoduodenal arteries supply the duodenum, head of the pancreas and bile duct. The entrance of the bile duct marks the junction of foregut and midgut, and is the meeting place of the arterial distributions of their respective arteries, coeliac and superior mesenteric.
One or two small supraduodenal arteries may arise from the common hepatic artery or its branches.
Venous drainage of the foregut
Right and left gastric, right and left gastroepiploic and the short gastric veins run with the corresponding arteries. All this blood reaches the liver via the portal vein (see p. 266) and, with the arterial blood of the hepatic artery, passes through the liver to be carried via the hepatic veins to the inferior vena cava.
The lower third of the oesophagus in the posterior mediastinum drains downwards by oesophageal veins, through the oesophageal opening in the diaphragm, to the left gastric vein. The oesophagus above this level drains into the azygos system of veins.
The left gastric vein runs to the left along the lesser curvature up to the oesophagus, then passes medially behind the peritoneum of the posterior wall of the lesser sac to join the portal vein at the upper border of the first part of the duodenum. The right gastric vein runs along the lesser curvature to the pylorus and empties into the portal vein. It receives the prepyloric vein which ascends in front of the pylorus.
The short gastric and left gastroepiploic veins run with the arteries through the gastrosplenic ligament and greater omentum to the hilum of the spleen, where they empty into the splenic vein.
The splenic vein begins in the hilum of the spleen by confluence of half a dozen tributaries from that organ. Having received the short gastric and left gastroepiploic veins, it passes to the right with the tail of the pancreas, below the splenic artery, in the splenorenal ligament and continues to the right, posterior to the body of the pancreas (Fig. 5.26), which it grooves. In its course it lies on the hilum of the left kidney, the left psoas muscle and left sympathetic trunk, the left crus of the diaphragm, the aorta and superior mesenteric artery and the inferior vena cava. It lies in front of the left renal vein along the upper border of that vessel. In front of the inferior vena cava it joins the superior mesenteric vein at a right angle to form the portal vein. It receives many tributaries from the tail, body, neck and head of the pancreas. As it lies in front of the left crus of the diaphragm it receives the inferior mesenteric vein from the hindgut.
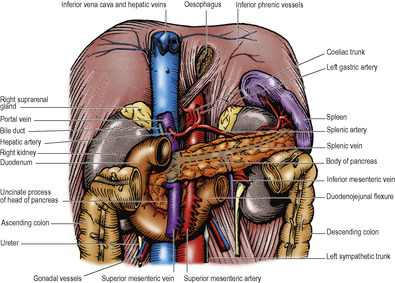
The right gastroepiploic vein runs to the right in the greater omentum and descends over the front of the pancreas to join the superior mesenteric vein at the lower border of the neck of the pancreas.
The superior pancreaticoduodenal vein ascends behind the head of the pancreas to join the portal vein at the upper border of the pancreas.
Blood supply of the midgut
Superior mesenteric artery
This is the artery of the midgut and supplies the gut from the entrance of the bile duct to a level just short of the splenic flexure of the colon. The artery arises from the front of the aorta a centimetre below the coeliac trunk, at the level of the lower border of L1 vertebra. It is directed steeply downwards behind the splenic vein and the body of the pancreas, with the superior mesenteric vein on its right side. It lies anterior to the left renal vein, the uncinate process of the pancreas and the third part of the duodenum, in that order from above downwards (Fig. 5.26). With its vein it enters the upper end of the mesentery of the small intestine and passes down to the right along the root of the mesentery (Fig. 5.23). Pressure of the superior mesenteric artery on the left renal vein may produce left-sided varicocele, and pressure on the duodenum may give symptoms of chronic duodenal ileus, particularly when the angle between the artery and the aorta is smaller than usual.
The inferior pancreaticoduodenal artery is its first branch, arising from the posterior surface; it may come off the first jejunal branch. It divides into anterior and posterior branches which run in the curve between the duodenum and the head of the pancreas, supply both, and anastomose with the terminal branches of the superior pancreaticoduodenal artery.
The jejunal and ileal branches arise from the left of the main trunk and pass down between the two layers of the mesentery. The pattern of anastomosing arcades from which vessels enter the gut wall is described with the jejunum and ileum (see p. 254).
The ileocolic artery (Fig. 5.23) arises from the right side of the superior mesenteric trunk low down in the base of the mesentery. It descends to the right iliac fossa and divides into superior and inferior branches. The superior branch runs up along the left side of the ascending colon to anastomose with the right colic artery. The inferior branch runs to the ileocolic junction, and gives off anterior and posterior caecal arteries, an appendicular artery, and an ileal branch which ascends to the left on the ileum to anastomose with the terminal branch of the superior mesenteric artery.
The right colic artery (Fig. 5.23) arises from the right side of the superior mesenteric artery, or in common with the ileocolic artery. It runs to the right across the right psoas muscle, gonadal vessels, ureter and genitofemoral nerve, and quadratus lumborum, just behind the peritoneal floor of the right infracolic compartment. It divides near the left side of the ascending colon into two branches. The descending branch runs down to anastomose with the superior branch of the ileocolic artery. The ascending branch runs up across the inferior pole of the right kidney to the hepatic flexure where it anastomoses with a branch of the middle colic artery.
The middle colic artery (Fig. 5.23) arises from the right side of the superior mesenteric artery, as the artery emerges at the lower border of the neck of the pancreas, and descends between the two leaves of the transverse mesocolon. It lies to the right of the midline and at the intestinal border of the transverse mesocolon it divides into right and left branches which run along the margin of the transverse colon. The right branch anastomoses with the ascending branch of the right colic artery. The left branch supplies the transverse colon almost to the splenic flexure (the distal part of the midgut) where it anastomoses with a branch of the left colic artery. As the middle colic lies to the right of the midline it leaves a large avascular window to its left in the transverse mesocolon (Fig. 5.23). This window is the site of election for surgical access to the lesser sac and the posterior wall of the stomach.
Venous drainage of the midgut
Each branch of the superior mesenteric artery is accompanied by a vein. All these veins flow into the superior mesenteric vein, a large trunk which lies to the right of the artery. It crosses the third part of the duodenum and the uncinate process of the pancreas. Behind the neck of the pancreas it is joined by the splenic vein to form the portal vein. This continues upwards behind the first part of the duodenum. The superior mesenteric and portal veins represent a single continuing venous trunk, named portal vein above, and superior mesenteric vein below, the level of entry of the splenic vein (Fig. 5.26).
Blood supply of the hindgut
Inferior mesenteric artery
This is the artery of the hindgut; its area of supply extends as far as the upper third of the anal canal, i.e. to the level of the pectinate (dentate) line (see p. 315). The inferior mesenteric artery arises from the front of the aorta behind the inferior border of the third part of the duodenum, opposite L3 vertebra, at the level of the umbilicus, 3 or 4cm above the aortic bifurcation. It is smaller than the superior mesenteric artery. It runs obliquely down to the pelvic brim, immediately beneath the peritoneal floor of the left infracolic compartment. It gives off the left colic and sigmoid arteries. It crosses the pelvic brim at the bifurcation of the left common iliac vessels over the sacroiliac joint, at which point it converges towards the ureter, with the inferior mesenteric vein lying between them, at the apex of the Λ-shaped attachment of the sigmoid mesocolon. Its branches cross to the left in front of the ureter and the other structures in the posterior abdominal wall of the left infracolic compartment. Beyond the pelvic brim it continues in the root of the sigmoid mesocolon as the superior rectal artery (see p. 294).
The left colic artery leaves the trunk and passes upwards and to the left behind the peritoneum. After a short course it divides into an ascending and a descending branch. The ascending branch continues laterally and upwards, crossing the left psoas muscle, gonadal vessels, ureter and genitofemoral nerve, and quadratus lumborum. It is crossed anteriorly by the inferior mesenteric vein. The descending branch passes laterally and downwards. The branches of these two arteries anastomose with each other as well as (above) with the left branch of the middle colic artery and (below) with the highest sigmoid artery, thus contributing to an anastomotic channel along the inner margin of the colon (Fig. 5.24 and p. 258).
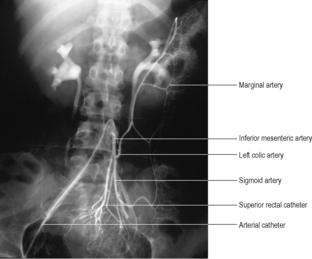 |
| Figure 5.24 |
The sigmoid arteries are two to four branches which pass between the layers of the sigmoid mesocolon, in which they form anastomosing loops. The last sigmoid branch anastomoses with the first branch of the superior rectal artery.
Venous drainage of the hindgut
The superior rectal vein runs up in the root of the sigmoid mesocolon, on the left of the superior rectal artery, to the pelvic brim, above which it is named the inferior mesenteric vein. This receives tributaries identical with the branches of the inferior mesenteric artery. The vein itself runs vertically upwards well to the left of the artery, beneath the peritoneal floor of the left infracolic compartment. It lies on the left psoas muscle, in front of the gonadal vessels, ureter and genitofemoral nerve. At the upper limit of the left infracolic compartment, just below the attachment of the transverse mesocolon, it lies to the left of the duodenojejunal flexure. Here it curves towards the right and often raises up a ridge of peritoneum. This ridge may be excavated by a small recess of peritoneum, thus making a shallow cave, the paraduodenal recess (see p. 254).
The inferior mesenteric vein now passes behind the lower border of the body of the pancreas, in front of the left renal vein, and joins the splenic vein. Occasionally it curves to the right more sharply, and passes behind the pancreas, below and parallel with the splenic vein, in front of the superior mesenteric artery, to open directly into the superior mesenteric vein.
Lymph drainage of the gastrointestinal tract
From the whole length of the gastrointestinal tract the lymph vessels pass back along the arteries to lymph nodes that lie in front of the aorta at the origins of the gut arteries (Fig. 5.17). These comprise the coeliac, superior mesenteric and inferior mesenteric groups of lymph nodes. They drain into each other from below upwards, the coeliac group itself draining by two or three lymph channels into the cisterna chyli.
These preaortic lymph nodes are the last in a series of lymph node filters that lie between the mucous membrane of the gut and the cisterna chyli. The first filtering mechanism consists of isolated lymphoid follicles which lie in the mucous membrane of the alimentary canal from mouth to anus. They are not numerous in the oesophagus, but are numerous in the stomach, and become increasingly so along the small intestine (the MALT, see p. 9). In the lower reaches of the ileum they become aggregated together into Peyer’s patches visible through the muscular wall. These lie on the antimesenteric border of the ileum and are oval in shape, with their long axes lying longitudinally along the ileum. In the large intestine the lymphoid follicles in the mucous membrane are numerous, but isolated from each other. In the appendix they are aggregated as in a tonsil.
Lymph vessels pass from the follicles in the mucous membrane through the muscle wall of the gut to nearby nodes, which also receive lymph from minute nodules on the serosal surface of the gut. The various groups of nodes are considered with the individual organs and have particular significance for the stomach and large intestine in view of the prevalence of carcinoma at these sites. However, it is convenient to note here that the small and large intestines have a common pattern of three groups of nodes. The first group lies in the peritoneum adjacent to the margin of the gut, the mural nodes in the mesentery of the small intestine, and the paracolic nodes of the large intestine. The second group of intermediate nodes lies along the main blood vessels of supply, and the third are the preaortic nodes at the origins of the coeliac and the superior and inferior mesenteric arteries. The large intestine has some additional nodes which lie on the external surface of the gut wall (and occasionally within appendices epiploicae); these are the epicolic nodes.
Nerve supply of the gastrointestinal tract
All parts of the gut and its derivatives are innervated by parasympathetic and sympathetic nerves, which travel together along the gut arteries to reach their destination. Most come from the coeliac plexus and connected plexuses on the abdominal aorta (see p. 281), but parasympathetic fibres to the hindgut come from the inferior hypogastric plexus in the pelvis (see p. 311).
From the middle third of the oesophagus to the rectum, nerve cells and fibres that supply muscle, blood vessels and glands in the alimentary tract are concentrated in two plexuses. The myenteric plexus (of Auerbach) is situated between the two muscle layers of the gut and the submucous plexus (of Meissner) is in the submucosa. Collectively the plexuses form the enteric nervous system. The system receives postganglionic sympathetic (inhibitory) and preganglionic parasympathetic (excitatory) fibres, but is unique in being able to function without these extrinsic efferent supplies, which do not pass directly to the gut muscle; the enteric system always intervenes. Afferent fibres connect with the spinal cord (by the sympathetic trunks) and with the brainstem (by the vagus). Pain impulses are transmitted by sympathetic and parasympathetic fibres, while impulses mediating sensations of distension pass in parasympathetic fibres.
Part six. Gastrointestinal tract
General structural features
The alimentary canal has its embryological origin in the yolk sac. It is lined by epithelium derived from the endoderm, and outgrowths from this epithelium (liver and pancreas) are thus endodermal in origin.
Macroscopically the alimentary canal is a tube of muscle lined with mucous membrane and covered, for the most part, with peritoneum.
The muscular wall of the alimentary canal, from the upper end of the oesophagus to the lower end of the rectum, is in two separate layers, an inner circular and an outer longitudinal. This arrangement is characteristic of all tubes that empty by orderly peristalsis and not by a mass contraction. In reality the muscle is in two spiral layers. The inner layer forms a closewound spiral and is the circular layer. The outer layer spirals in such gradual fashion that its fibres are virtually longitudinal, the longitudinal layer. The latter is mostly separated into three discrete bundles, the taeniae coli, in the colon. An innermost third layer of fibres arranged in an oblique manner reinforces the body of the stomach.
The two layers of the upper third of the oesophagus (cervical and superior mediastinal parts) consist of striated muscle. In the lower two-thirds of the oesophagus this is gradually replaced by visceral muscle, which constitutes the two layers of the remainder of the alimentary canal.
The mucous membrane of the alimentary tube consists of three components: epithelium, an underlying connective tissue layer or lamina propria, and a thin layer of smooth muscle, the muscularis mucosae.
A variety of neuroendocrine cells are scattered in the mucosa of the alimentary tract from oesophagus to anal canal. The number of these cells progressively decreases in an anal direction. They all produce peptides and/or amines, which are active as hormones or neurotransmitters. They are a part of the amine precursor uptake and decarboxylation (APUD) cell series, and they modulate autonomic activity as well as each other. The substances produced by these cells include gastrin, vasoactive intestinal polypeptide (VIP), cholecystokinin (CCK), motilin, secretin, somatostatin, substance P (SP), serotonin and endorphins.
Oesophagus
The epithelium is stratified squamous non-keratinizing, like that of most of the pharynx and mouth. The muscularis mucosae, which is absent from the uppermost part, is characteristically thicker in the oesophagus than in any other part of the alimentary tract. Small groups of mucus-secreting glands may be present at the upper and lower ends, in both the mucosa and submucosa.
Stomach
At the gastro-oesophageal junction (cardia), or a few centimetres above it, there is an abrupt change along an irregular line from greyish-pink stratified to reddish-pink single-layered columnar epithelium which continues all the way to the anal canal, but with differing cell types and configuration in the different organs. Throughout, the epithelium is not a flat layer but dips down into the connective tissue lamina propria to form myriads of glands. In the main part of the stomach, the body, the mucus-secreting surface cells dip down to form the gastric pits which in turn continue downwards as the straight test-tube-like glands whose cells include the peptic and parietal cells secreting pepsin and hydrochloric acid respectively. The parietal cells also secrete intrinsic factor, which is necessary for the intestinal absorption of vitamin B12. For about a 1cm ring at the cardia, the glands are shorter and the cells all mucus-secreting. In the pyloric region the glands are like coiled test-tubes and again the cells are all mucus-secreting. This part of the stomach contains most of the gastrin-producing endocrine G cells, as well as D cells which secrete somatostatin. Chromaffin endocrine cells in both body and pyloric regions produce serotonin and endorphin.
Small intestine
In the whole length of the small intestine, the columnar epithelium not only dips down to form glands (the crypts of Lieberkühn) but is also thrown up between the gland openings into villi, which consist of finger-like or leaf-like connective tissue cores covered by epithelium. Some of the villous cells are mucus-secreting goblet cells; others are the absorbing cells or enterocytes. Both types are derived from progenitors in the crypts and are constantly being shed and renewed, every few days. (So are other alimentary epithelial cells but at slower rates.) At the bases of the crypts are the granular Paneth cells which secrete lysozyme. Scattered among the other crypt cells are various neuroendocrine cells responsible for the production of intestinal hormones, including secretin, somatostatin and CCK, as well as chromaffin cells.
The duodenum is distinguished from the rest of the small intestine in having mucus-secreting glands (of Brunner) in the submucosa. In the terminal ileum there are groups of lymphoid follicles in the mucosa forming Peyer’s patches, in contrast to single follicles which can be found throughout the alimentary canal from mouth to anus.
Large intestine
In the large intestine there are no villi, only glands (crypts) containing a high proportion of goblet cells. In the appendix the glands are rather shallower and less closely packed than in the rest of the large intestine, and there are numerous lymphoid follicles in the mucosa and submucosa.
In the upper part of the anal canal, the columnar epithelium gives place to the stratified squamous type, but here the junction is not as clear cut as the change at the cardia (see above).
Oesophagus
The abdominal oesophagus, about 1 to 2cm in length, turns forwards and to the left immediately below the diaphragmatic opening and grooves the posterior surface of the left lobe of the liver. The anterior and posterior vagal trunks are related to the respective oesophageal surfaces here (see p. 250). It is covered on its front and left side by peritoneum which passes from it on the right to the diaphragm as the uppermost part of the lesser omentum, and on the left forms the uppermost part of the greater omentum. The posterior wall of the oesophagus is rather shorter than the anterior, for the orifice in the diaphragm lies very nearly vertical. The left inferior phrenic artery lies behind the oesophagus.
The oesophagus enters the stomach at the cardiac orifice. The right margin is continuous with the lesser curvature, while the left margin makes an acute angle with the gastric fundus, the cardiac notch. The fibres of the right crus that pass to the left of the diaphragmatic opening form a sling around the abdominal oesophagus. Various factors contribute towards guarding against the reflux of gastric contents; these include the contraction of these right crus fibres, the angle of entry of the oesophagus into the stomach, the longitudinal folds of the oesophageal mucosa, a high pressure zone in the lower 3cm of oesophagus and the effect of positive intra-abdominal pressure on the abdominal segment of the oesophagus.
Stomach
The stomach is the most dilated part of the alimentary tract, interposed between the oesophagus and duodenum in the upper part of the abdominal cavity. Lying mainly in the left hypochondrial, epigastric and umbilical regions, much of it is under cover of the lower ribs. It is a muscular bag, relatively fixed at both ends but otherwise subject to great variations in size depending on the volume of its contents. The gastro-oesophageal junction is the cardia, the most fixed part of the organ, and lies 2.5cm to the left of the midline at the level of T10 vertebra, behind the seventh left costal cartilage. It is 40cm from the incisor teeth. The gastroduodenal junction is the pylorus. In the recumbent position with the stomach empty this is usually a little to the right of the midline at the level of L1 vertebra but may be considerably lower.
The main parts of the stomach are the fundus, body and pyloric part, with the greater and lesser curvatures forming the left and right borders respectively. The stomach is completely invested by peritoneum, which passes in a double layer from the lesser curvature to the liver as the lesser omentum, and hangs down from the fundus and greater curvature as the greater omentum.
The fundus is the part which projects upwards above the level of the cardia, and is in contact with the left dome of the diaphragm. It is usually full of swallowed air.
The largest part of the stomach is the body, extending from the fundus to the notch, the angular incisure, on the lower part of the lesser curvature. Variable amounts of the body will be above and below the costal margin, in contact with the diaphragm and anterior abdominal wall. Likewise the lowest part of the greater curvature may be above or below umbilical level.
The pyloric part extends from the angular notch to the gastroduodenal junction, and consists of the proximal pyloric antrum which narrows distally as the pyloric canal (Fig. 5.25). The circular muscle of the distal end of the canal is palpably thickened to form the pyloric sphincter, whose position is indicated on the anterior surface by the prepyloric vein (of Mayo). The pyloric canal lies on the head and neck of the pancreas.
Behind the stomach are a group of structures comprising the stomach bed (Fig. 5.26). The posterior wall of the stomach is covered by peritoneum of the anterior wall of the lesser sac, and the bed is covered by the lesser sac’s posterior wall. Apart from the left crus and dome of the diaphragm the bed consists of the splenic artery, body of pancreas, transverse mesocolon, upper part of left kidney, left suprarenal gland, spleen and left colic flexure. To the right of the lesser curvature in the midline lies the aorta with the coeliac trunk, the coeliac plexus and ganglia, and coeliac lymph nodes.
Blood supply
The stomach is supplied by branches from the coeliac trunk. Along the lesser curvature between the two layers of the lesser omentum the left gastric artery anastomoses with the right gastric; these arteries may be double. The fundus and upper left part of the greater curvature receive about six short gastric arteries from the splenic artery in the gastroplenic ligament. The rest of the curvature is supplied by the left and right gastroepiploic vessels, which run between the two layers of the greater omentum and anastomose with each other. The right gastroepiploic artery is closer to the greater curvature than the left gastroepiploic artery. During partial gastrectomy the greater omentum is divided below the right and above the left gastroepiploic arteries; the blood supply to the omentum usually remains intact as the larger omental branches of the left gastroepiploic artery are preserved. A branch from the splenic artery as it runs along the upper border of the pancreas, the posterior gastric artery, may also supply the stomach.
Veins of the same name accompany the arteries and drain into the portal vein itself or its splenic and superior mesenteric tributaries. The prepyloric vein (unaccompanied by an artery) drains into the right gastric vein.
Lymph drainage
All lymph from the stomach eventually reaches coeliac nodes after passing through various outlying groups (Fig. 5.27A). Lymph vessels anastomose freely in the stomach wall, but there are valves in the vessels that direct lymph in such a way that a line drawn parallel to the greater curvature and two-thirds of the way down the anterior surface indicates a ‘watershed’ (Fig. 5.27B). From the largest zone above and to the right of this line, lymph passes to left and right gastric nodes along the left and right gastric arteries. From the upper left quadrant lymph flows via left gastroepiploic nodes and directly to pancreaticosplenic nodes at the splenic hilum, and at the upper border and posterior surface of the pancreas, accompanying the splenic artery. From the rest of the greater curvature region of the stomach lymph reaches nodes along the right gastroepiploic vessels, which drain to subpyloric nodes near the gastroduodenal artery. The pyloric part of the stomach drains to hepatic nodes in the porta hepatis and to subpyloric and right gastric nodes. In cases of gastric carcinoma, the left supraclavicular nodes may rarely become palpably involved (Troisier’s sign), presumably by spread along the thoracic duct.
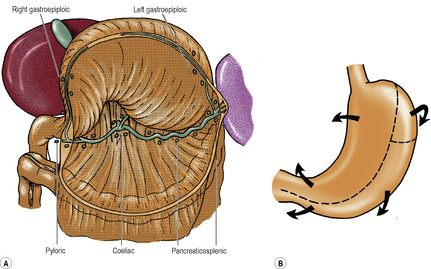
Nerve supply
Sympathetic fibres (which are vasomotor and also cause pyloric sphincter contraction) accompanied by afferent (pain) fibres run with the various arterial branches to the stomach. The parasympathetic supply from the vagi controls motility and secretion (although 90% of vagal fibres below the diaphragm are afferent for reflex activities, not pain). At the oesophageal opening in the diaphragm the anterior vagal trunk (comprising mainly left vagal fibres from the oesophageal plexus in the posterior mediastinum) lies in contact with the anterior oesophageal wall (Fig. 5.28), usually nearer its right margin; sometimes this trunk is double or triple. Each anterior trunk gives off one or two hepatic branches, which run in the upper part of the lesser omentum to join the plexus on the hepatic artery and portal vein, and then turn down in the anterior wall of the epiploic foramen to reach the pylorus. The anterior vagus also gives off several gastric branches which supply the fundus and body, and one large branch (greater anterior gastric nerve or anterior nerve of Latarget) which runs down in the lesser omentum near the lesser curvature with the left gastric artery, and subdivides in the manner of a crow’s foot to supply the antrum and pyloric sphincter. The posterior vagal trunk (comprising mainly right vagal fibres) lies in loose tissue a little behind and to the right, not in contact with the posterior surface of the oesophagus (Fig. 5.28). It gives coeliac branches that run backwards along the left gastric artery to the coeliac plexus. It also gives gastric branches to the fundus and body, and a large branch (greater posterior gastric nerve or posterior nerve of Latarget) which runs in the lesser omentum behind the anterior trunk to reach the antrum but not the pyloric sphincter.
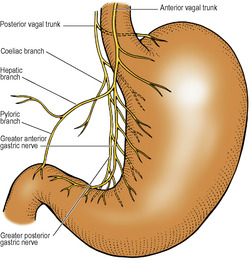
Vagotomy. Truncal vagotomy involves cutting the trunks at the level of the abdominal oesophagus. In selective vagotomy the vagal trunks are cut distal to the hepatic branch of the anterior vagus and the coeliac branches of the posterior vagus. Although effective in diminishing gastric secretion, truncal and selective vagotomy are often accompanied by gastric stasis, so that an antral drainage procedure is required. Highly selective vagotomy (parietal cell vagotomy) attempts to avoid stasis by cutting only the branches to the fundus and body, leaving the antral and pyloric branches intact. Arterial branches run into the lesser curvature transversely but nerve branches approach it obliquely. Ligating vessels will inevitably sever some nerve branches but not all, as not all nerves accompany vessels closely, and any individual nerves that can be identified must be cut also.
Structure
The main histological features are referred to on page 248. Although the angular notch is usually taken as the dividing line between the body and pyloric parts of the stomach, this does not necessarily indicate exactly where the body-type of mucosa with its parietal (acid-secreting) cells gives way to the pyloric-type with its concentration of G cells (producing gastrin). The outer longitudinal and inner circular muscle coats completely invest the stomach. They are reinforced by an incomplete innermost oblique muscle layer; its fibres loop over the fundus, being thickest at the notch between oesophagus and stomach, helping to maintain the angle here.
Small intestine
The small intestine consists of the duodenum, jejunum and ileum, although clinically the term small intestine often excludes the duodenum.
Duodenum
The duodenum is a C-shaped tube lying in front of, and to the right of the inferior vena cava and aorta (Fig. 5.26). The first 2.5cm are contained between the peritoneum of the lesser and greater omenta, but the remainder is retroperitoneal. It is divided into four parts, superior, descending, horizontal and ascending, or more simply first, second, third and fourth. The total length is 25cm (10 in), the lengths of the parts being easily remembered in inches as 2, 3, 4 and 1 but less conveniently in centimetres (5, 7.5, 10 and 2.5).
The duodenum makes its C-shaped loop round the head of the pancreas, which is opposite the body of L2 vertebra; so the first part may be said to lie at the level of L1, the second on the right side of L2, the third crosses in front of L3, and the fourth is on the left of L2 vertebrae.
The first part of the duodenum runs to the right, upwards and backwards from the pylorus; a foreshortened view is consequently obtained in anteroposterior radiographs (Fig. 5.25). The first 2.5cm (i.e. the duodenal cap, see below) lies between the peritoneal folds of the greater and lesser omenta; it forms the lowermost boundary of the opening into the lesser sac (Fig. 5.35A). It lies in front of the gastroduodenal artery, bile duct, and portal vein, and behind these structures lies the inferior vena cava. The gallbladder is anterior to the duodenal cap. The next 2.5cm passes backwards and upwards on the upper part of the head of the pancreas to the medial border of the right kidney. It is covered in front with peritoneum, and the inferior surface of the right lobe of the liver lies over this peritoneum. Its posterior surface is bare of peritoneum.
The second part of the duodenum curves downwards over the hilum of the right kidney (Fig. 5.26). It is covered in front with peritoneum and crossed by the attachment of the transverse mesocolon, so that its upper half lies in the supracolic compartment to the left of the hepatorenal pouch (in contact with the liver) and its lower half lies in the right infracolic compartment medial to the inferior pole of the right kidney (in contact with coils of jejunum). It lies alongside the head of the pancreas, approximately at the level of L2 vertebra.
Its posteromedial wall receives the common opening of the bile duct and main pancreatic duct at the hepatopancreatic ampulla (of Vater), which opens on the summit of the major duodenal papilla, halfway along the second part, 10cm from the pylorus. It is overlapped by a semilunar flap of mucous membrane. Two centimetres proximal is the small opening of the accessory pancreatic duct (on the minor duodenal papilla).
The third part of the duodenum curves forwards from the right paravertebral gutter over the slope of the right psoas muscle (gonadal vessels and ureter intervening) and passes over the forwardly projecting inferior vena cava and aorta to reach the left psoas muscle (Fig. 5.26). As its inferior border crosses the aorta it lies on the commencement of the inferior mesenteric artery. Its upper border hugs the lower border of the pancreas. It is covered by the peritoneum of the posterior abdominal wall just below the transverse mesocolon. It is crossed by the superior mesenteric vessels and by the leaves of the commencement of the mesentery of the small intestine sloping down from the duodenojejunal flexure. It lies, therefore, in both right (mainly) and left infracolic compartments (Fig. 5.14). Its anterior surface is in contact with coils of jejunum.
The fourth part of the duodenum ascends to the left of the aorta, lying on the left psoas muscle and left lumbar sympathetic trunk, to reach the lower border of the body of the pancreas. It is covered in front by the peritoneal floor of the left infracolic compartment and by coils of jejunum. It breaks free from the peritoneum that has plastered it down to the posterior abdominal wall and curves forwards and to the right as the duodenojejunal flexure. This pulls up a double sheet of peritoneum from the posterior abdominal wall, the mesentery of the small intestine, which slopes down to the right across the third part of the duodenum and posterior abdominal wall (Fig. 5.29).
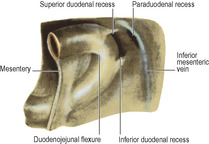
The duodenojejunal flexure is fixed to the left psoas fascia by fibrous tissue and may be further supported by the suspensory muscle of the duodenum (muscle or ligament of Treitz). This is a thin band of connective tissue which may contain muscle—skeletal muscle fibres that run from the right crus of the diaphragm to connective tissue around the coeliac trunk and smooth muscle fibres that run from there, behind the pancreas and in front of the left venal vein, to the muscle coat of the flexure.
Internally the mucous membrane of most of the duodenum, like the rest of the small intestine, is thrown into numerous circular folds (plicae circulares or valvulae conniventes). But the walls of the first 2.5cm are smooth, hence the smooth outline of the full shadow of barium in the ‘duodenal cap’ at radiographic examination (Fig. 5.25). From the duodenal cap onwards the plicae break up the barium and its shadow.
Paraduodenal recesses. To the left of the duodenojejunal flexure certain peritoneal folds may cover recesses or fossae. The paraduodenal recess proper (Fig. 5.29) lies behind a fold raised by the upper end of the inferior mesenteric vein; an incarcerated internal hernia in this fossa may obstruct and thrombose the vein, and there is danger of dividing the vein if the peritoneum has to be divided at operation to free the hernia. Horizontal folds of peritoneum may cover superior and inferior duodenal recesses and a retroduodenal recess may be found behind the flexure. The mouths of these four recesses or fossae all face towards each other.
Blood supply
The duodenum is supplied by the superior and inferior pancreaticoduodenal arteries, but the first 2cm, the usual site of ulceration, receives blood from the hepatic, gastroduodenal, supraduodenal, right gastric and right gastroepiploic arteries. Venous drainage is to tributaries of the superior mesenteric and portal veins.
Lymph drainage
Duodenal lymph drains by channels that accompany the superior and inferior pancreaticoduodenal vessels to coeliac and superior mesenteric nodes.
Jejunum and ileum
The jejunum is wider-bored and thicker-walled than the ileum. The thick wall of the jejunum feels double (the mucous membrane can be felt through the muscle wall, ‘a shirt sleeve felt through a coat sleeve’); the thin wall of the ileum feels single.
The lower reaches of the ileum are distinguished by the presence on the antimesenteric border of elongated whitish plaques in the mucous membrane, usually but not always visible through the muscle wall. These are the aggregated lymphoid follicles (Peyer’s patches). The jejunum lies coiled in the upper part of the infracolic compartment, the ileum in the lower part thereof and in the pelvis.
The jejunum and ileum together lie in the free margin of the mesentery. Total length varies greatly, from about 4 to 6 metres. The jejunum constitutes rather less than half the total length, say two-fifths, allowing three-fifths for the ileum.
An ileal (Meckel’s) diverticulum is present in 2% of individuals, 60cm (2ft) from the caecum, and is 5cm (2 in) long, according to a popular mnemonic but the length of the diverticulum is variable and its site may be more proximal. Its blind end may contain gastric mucosa or liver or pancreatic tissue; ulceration and perforation of the tip can occur. It represents the intestinal end of the vitellointestinal duct, and its apex may be adherent to the umbilicus or connected thereto by a fibrous cord, a further remnant of the duct.
Blood supply
Numerous jejunal and ileal branches arise from the left side of the superior mesenteric artery and enter the mesentery by passing between the two layers of the root. The jejunal branches join each other in a series of anastomosing loops to form arterial arcades: single for the upper jejunum and double lower down. From the arcades, straight arteries pass to the mesenteric border of the gut (Fig. 5.30). These vessels are long and close together, forming high narrow ‘windows’ in the intestinal border of the mesentery, visible because of the scantiness of mesenteric fat here. The straight vessels pass to one or other side of the jejunum and sink into its wall. Occlusion of a straight artery may lead to infarction of the segment supplied because these are end arteries, but occlusion of arcade vessels is usually without effect due to their numerous anastomotic connections.
The ileal arteries are similar but form a larger series of arcades—three to five, the most distal lying near the ileal wall so that the straight vessels branching off the arcades are shorter. There is more fat in this part of the mesentery, so the windows characteristic of the jejunal part are not seen; this is a useful feature in identifying loops of the bowel.
The end of the superior mesenteric artery itself supplies the region of the ileal diverticulum (if present), and anastomoses with the arcades and with the ileocolic branch to supply the terminal ileum.
The veins all correspond to the arteries and thus drain to the superior mesenteric.
Lymph drainage
Jejunal and ileal lymph drains to superior mesenteric nodes via mural and intermediate nodes in the mesentery.
Nerve supply
Autonomic nerves reach the wall of the small intestine with its blood vessels. The parasympathetic vagal supply augments peristaltic activity and intestinal secretion. There are many afferent fibres whose function is uncertain. The sympathetic supply, which is vasoconstrictor and normally inhibits peristalsis, is from the lateral horn cells of spinal segments T9 and 10. Pain impulses use sympathetic pathways mainly and small intestinal pain is usually felt in the umbilical region of the abdomen.
Large intestine
The large intestine consists of the caecum with the (vermiform) appendix, the ascending, transverse, descending and sigmoid parts of the colon, the rectum and the anal canal.
Caecum and appendix
Caecum
This blind pouch of the large intestine projects downwards from the commencement of the ascending colon, below the ileocaecal junction (Fig. 5.31A). It is usually completely covered by peritoneum, which is reflected downwards to the floor of the right iliac fossa, and the retrocaecal peritoneal space may be shallow or deep, according to the distance of this reflection from the lower end of the caecum. Often there are two peritoneal folds from either side of the posterior wall of the caecum, forming between them the retrocaecal recess in which the appendix may lie. As in the rest of the colon, the longitudinal muscle of the caecum is concentrated into three flat bands, the taeniae coli, within which is the circular muscle layer of the sacculated wall. The taeniae lie one anterior, one posteromedial and one posterolateral. All three converge on the base of the appendix, to which they are a useful guide.
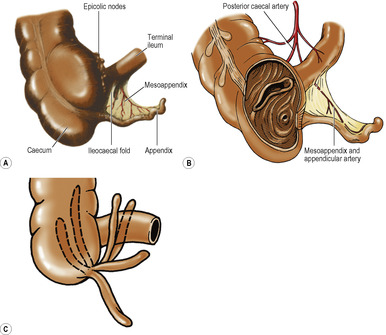
Internally the ileocaecal junction is guarded by the ileocaecal valve (Fig. 5.31B), whose almost transverse lips may help to prevent some reflux into the ileum, but any possible sphincteric action is poor.
In the infant the caecum is conical and the appendix extends downwards from its apex. The lateral wall outgrows the medial wall and bulges down below the base of the appendix in the adult; the base of the appendix thus comes to lie in the posteromedial wall of the caecum above its lower end and the three taeniae coverage to this point. The endoscopic view of the site of convergence has a trefoil appearance (Fig. 5.31D).
The caecum lies on the peritoneal floor of the right iliac fossa, over the iliacus and psoas fasciae and the femoral and lateral femoral cutaneous nerves. Its lower end lies at the pelvic brim. When distended its anterior surface touches the parietal peritoneum of the anterior abdominal wall; when collapsed, coils of ileum lie between the two.
Blood supply. Branches of the anterior and posterior caecal arteries (branches of the ileocolic artery) fan out over the respective surfaces of the caecum. The posterior caecal artery is larger and gives a branch to the base of the appendix. There are corresponding veins.
Lymph drainage. Lymph passes to nodes associated with the ileocolic artery.
Appendix
The vermiform (worm-shaped) appendix is a blind-ending tube (Fig. 5.31) varying in length (commonly about 6–9cm), which opens into the posteromedial wall of the caecum 2cm below the ileocaecal valve (Fig. 5.31B). On the surface of the abdomen this point (McBurney’s) lies one-third of the way up the oblique line that joins the right anterior superior iliac spine to the umbilicus. While the position of its base is constant in relation to the caecum, the appendix itself may lie in a variety of positions (Fig. 5.31C). The most common, as found at operation, is the retrocaecal position, with the pelvic position next in order of frequency.
The three taeniae of the caecum merge into a complete longitudinal muscle layer for the appendix. The submucosa contains many lymphoid masses and the lumen is thereby irregularly narrowed. This lumen is wider in the young child and may be obliterated in old age.
The appendix has its own short mesentery, the mesoappendix, which is a triangular fold of peritoneum from the left (inferior) layer of the mesentery of the terminal ileum (Fig. 5.31A). A small fold of peritoneum extends from the terminal ileum to the front of the mesoappendix. This is the ileocaecal fold (or ‘bloodless fold of Treves’, although it sometimes contains blood vessels) and the space between it and the mesoappendix is the inferior ileocaecal recess. Another fold lies in front of the terminal ileum, between the base of the mesentery and the anterior wall of the caecum. This fold is raised up by the contained anterior caecal artery and is called the vascular fold of the caecum. The space behind it is the superior ileocaecal recess.
Blood supply. The appendicular artery is normally a branch of the inferior division of the ileocolic artery, which runs behind the terminal ileum to enter the mesoappendix. As it does so it gives off a recurrent branch which anastomoses with a branch of the posterior caecal artery; this may replace the appendicular artery. The appendicular artery runs first in the free margin of the mesoappendix and then close to the appendicular wall, where it may be thrombosed in appendicitis, leading to ischaemic necrosis and perhaps rupture of the appendix; there is no collateral circulation, for the appendicular artery is an end artery. There are corresponding veins.
Lymph drainage. As from the caecum, lymph passes to nodes associated with the ileocolic artery.
Appendicectomy. Exposure of the appendix during appendicectomy is through a McBurney or transverse muscle-splitting incision (see p. 232). If it is not immediately obvious, tracing any of the taeniae down over the caecal wall will lead to the base of the appendix.
Colon
Of the four parts of the colon, the transverse and sigmoid parts are suspended in mesenteries—the transverse mesocolon and sigmoid mesocolon respectively—but the ascending and descending colon are plastered on to the posterior abdominal wall so that they have posterior ‘bare areas’ devoid of peritoneum.
Ascending colon
This first part of the colon, about 15cm in length, extends upwards from the ileocaecal junction to the right colic (hepatic) flexure. The latter lies on the inferolateral part of the anterior surface of the right kidney, in contact with the inferior surface of the liver. The ascending colon lies on the iliac fascia and the anterior layer of the lumbar fascia. Its front and both sides possess a serous coat, which runs laterally into the paracolic gutter and medially into the right infracolic compartment. The original embryonic mesentery is retained in about 10% of adults.
The taeniae coli lie, in line with those of the caecum, anteriorly, posterolaterally and posteromedially. These consist of longitudinal muscle fibres and the circular muscle coat is exposed between them. The ascending colon is sacculated, due to the three taeniae being ‘too short’ for the bowel. If the taeniae are divided between the sacculations the latter can be drawn apart and the bowel wall flattened.
Small pouches of peritoneum, distended with fat, the appendices epiploicae, project in places from the serous coat. The blood vessels supplying them from the mucosa perforate the muscle wall. Mucous membrane may herniate through these vascular perforations, a condition known as diverticulosis. Diverticulitis is inflammation of these mucosal herniae.



