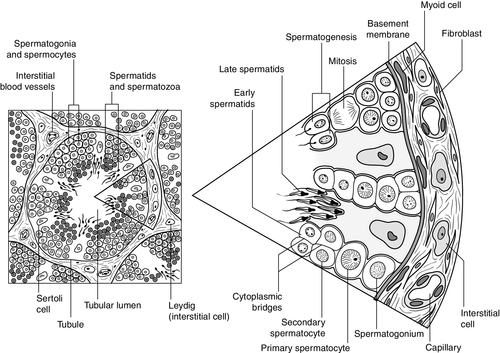
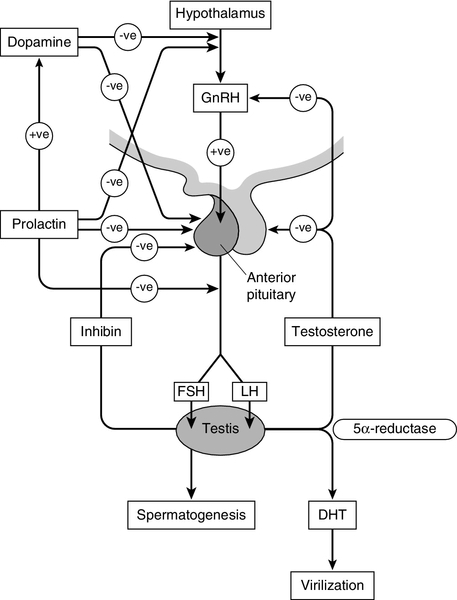
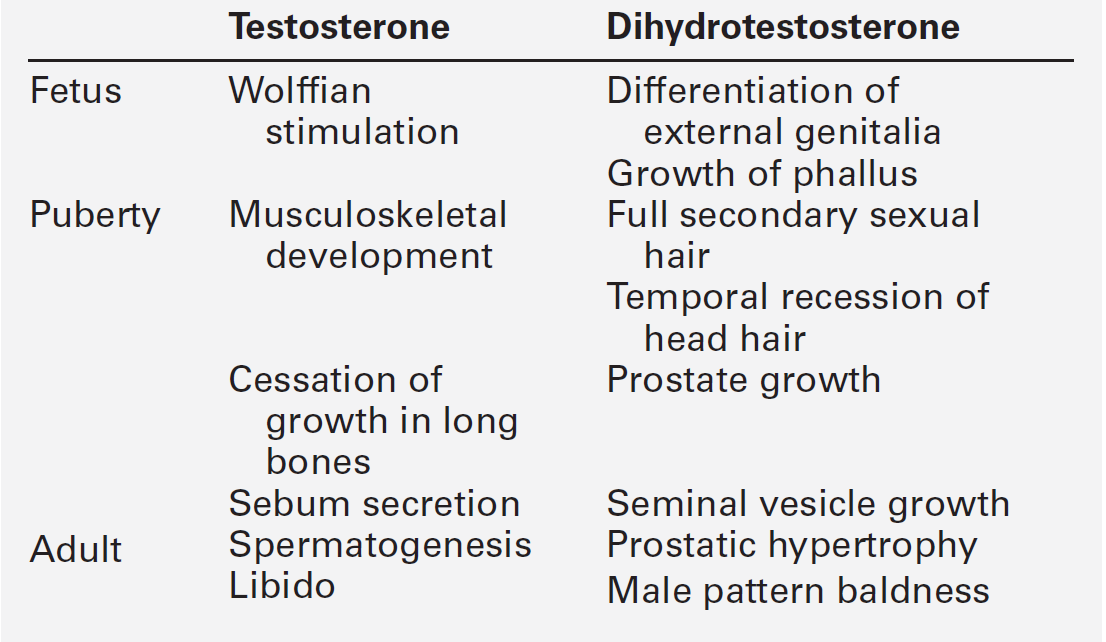
CHAPTER 23 CHAPTER OUTLINE Production and actions of testosterone Hypothalamo–pituitary control of testicular function Endocrine disrupting chemicals EVALUATION OF TESTICULAR FUNCTION Endocrine evaluation: hypothalamo–pituitary–gonadal axis Defective hormone synthesis and hormone receptor defects Treatment of erectile impotence Normal male reproductive function – the manufacturing of semen, the ability to create and sustain penile erection and to ejaculate – is dependent on the physiological integration of the hypothalamo–pituitary–testicular axis. Production of androgens, normal responses in the tissues on which these hormones act and intact neurological pathways required for ejaculation are crucial for full operation of this system, to make human reproduction possible. The testes have two distinct roles: androgen production and spermatogenesis. These are achieved by two main units – the interstitial cells and the seminiferous tubules – under the control of the hypothalamo–pituitary axis. Testosterone is the major androgen produced in the interstitial cells of the testes. Also known as Leydig cells, these are found in clumps between the seminiferous tubules, and their mass correlates positively with androgen production. Sertoli cells, along with germ cells, form the seminiferous tubules (Fig. 23.1) and secrete hormones that control embryological sexual differentiation. In addition, they support spermatogenesis by regulating the composition of the seminiferous tubular fluid, thereby providing an environment for meiotic germ cell development. FIGURE 23.1 The seminiferous tubules. At week eight of gestation, a human fetus has both Müllerian and Wolffian ducts, with the potential to develop into female and male genital tracts, respectively, and differing only by its sex chromosomes. The Y chromosome is a powerful sex determinant – it confers maleness to the extent that even XXXXY fetuses are phenotypically male at birth. (This topic is covered in more detail in Chapter 21.) Normal male sexual differentiation in utero depends on the presence of testosterone and other hormones produced by these embryological gonads. Leydig cells are stimulated by placental human chorionic gonadotrophin (hCG) to secrete testosterone from the ninth week of gestation, leading to development of the Wolffian ducts. Sertoli cells secrete anti-Müllerian hormone (AMH), which causes regression of the Müllerian ducts and oogonia (the precursors of primary oocytes). In the absence of AMH and testosterone, the Müllerian ducts differentiate into female internal genitalia. Lower concentrations of testosterone are maintained during late gestation by secretion of luteinizing hormone (LH) from the fetal pituitary. The presence of 5α-reductase, and the subsequent production of dihydrotestosterone (DHT) from testosterone, is required for development of the urogenital sinus. Testosterone secretion may continue for many months post partum before declining until the onset of puberty, when it will again be essential for sexual development. Testosterone secretion from Leydig cells follows stimulation by LH, which binds to specific membrane receptors. Luteinizing hormone activates adenylyl cyclase, resulting in formation of cAMP, which binds to protein kinase and in turn activates the enzyme 20,22-desmolase, which is responsible for conversion of cholesterol to pregnenolone (see Fig. 22.5, p. 438). The synthesis of testosterone is completed by four other enzymes (3β-hydroxysteroid dehydrogenase, 17α-hydroxylase, 17,20-desmolase and 17β-hydroxysteroid dehydrogenase). At the target organ, testosterone is transported to the nucleus, and gene activation results in the formation of specific messenger RNAs and proteins. Testosterone is usually converted to the active metabolite DHT by the enzyme 5α-reductase. In the adult, there is circadian variation in testosterone secretion with a peak in the early morning and a trough at about 18.00 h, and therefore blood samples for testosterone measurement should ideally be taken at 09.00 h. The biological actions of testosterone and DHT are summarized in Table 23.1. Gonadotrophin releasing hormone (GnRH) is secreted from the hypothalamus in a pulsatile fashion, with peaks occurring approximately every 90 min. It regulates the release of LH, which is crucial for the regulation of Leydig cell number and function, and hence production of testosterone, and follicle stimulating hormone (FSH), which stimulates Sertoli cell division and growth and is therefore responsible for testicular enlargement at puberty. Faster pulse frequencies favour LH release and slower ones, FSH. The pulsatility of GnRH secretion is important in the regulation of gonadotrophin secretion, as continuous exposure results in down-regulation of the GnRH receptor. Luteinizing hormone secretion is also highly pulsatile, which is reflected in measured plasma concentrations. Plasma concentrations of FSH are less variable, presumably as a result of slower metabolic clearance. The gonadotrophins are, in turn, regulated by a series of feedback mechanisms (Fig. 23.2). Testosterone inhibits LH and, to a lesser extent, FSH secretion from the anterior pituitary. Oestradiol, which is formed in the male through metabolism of androgens by aromatase found in adipose tissue, skin, kidneys and brain, in addition to Leydig cells, also inhibits gonadotrophin secretion by the pituitary. Opioids and prolactin reduce the pulsatile activity of GnRH and thus decrease FSH and LH secretion; prolactin also inhibits the direct action of gonadotrophins on the testes. FIGURE 23.2 Hypothalamo–pituitary control of testicular function. Follicle stimulating hormone is also regulated by inhibin B, which is produced by Sertoli cells and inhibits FSH production to regulate spermatogenesis. Conversely, activin works to enhance FSH action in the gonads, increasing spermatogenesis. Both are secreted in the seminal fluid and are thought to work locally in addition to centrally on the pituitary. Inhibin B can be used as a marker for male factor infertility as concentrations directly reflect the function of the Sertoli cells: plasma inhibin B concentrations are significantly higher in fertile men than in infertile men. Gonadotrophin secretion is affected by a diverse range of regulators, including stress and nutritional status. Recent studies have shown that low concentrations of leptin, an adipose-derived hormone, or decreased sensitivity to leptin, such as occurs in obese individuals, are associated with decreased gonadotrophin concentrations and decreased fertility. This is mediated via hypothalamic kisspeptin neurons rather than directly via GnRH neurons. Kisspeptin is a neurohormone thought to have a key role in puberty, although this is yet to be clearly defined. Patients with mutations in the kisspeptin gene fail to go through puberty owing to hypogonadotropic hypogonadism. In animal models, treatment with kisspeptin can restore GnRH pulsatility and gonadotrophin secretion. Testicular cancer is the commonest malignancy of men aged 20–39. Histologically, it is commonly a germ cell malignancy – either a seminoma or, more often, a teratoma. Patients are likely to present with a testicular mass, and as this cancer is relatively easily detectable with regular self-inspection/examination, and has a high cure rate if detected prior to metastatic spread, there have been several public campaigns to raise awareness amongst young men. Approximately 80% of testicular teratomas produce hCG or α-fetoprotein (AFP) (see Chapter 42). Human chorionic gonadotrophin is very similar to LH in structure. It stimulates the LH receptor in the testis eventually resulting in increased secretion of oestradiol, and hence gynaecomastia. Some rarer testicular malignancies, such as Sertoli cell tumours, can also secrete oestradiol directly. Human chorionic gonadotrophin can interact with the thyroid hormone axis as an agonist of the thyroid stimulating hormone (TSH) receptor, to cause hyperthyroidism. Endocrine disruptors are environmental substances that interfere with normal hormonal systems. They are very diverse and can be plastics, chemicals, pesticides and fungicides. They are often able to interact with steroid hormone receptors and can act as androgens and anti-androgens. Exposure may be occupational, but also through water supply, soil and food. There may be a lag between exposure and clinical presentation and there may be trans-generational effects if mutations or modifications of gene expression occur. The developmental stage at which a person is exposed may be crucial. The observations of a declining sperm count, and increasing rates of testicular cancer throughout the last century prompted researchers to ask if these epidemiological trends could be explained by endocrine disruptors. In animal models, exposure to phthalates is significantly related to oligospermia, and epidemiological evidence suggests this may be the case for humans. Endocrine disruptors have also been linked to cryptorchidism, hypospadias, early or delayed puberty and prostatic hyperplasia. More detail on this topic is beyond the scope of this chapter, but the interested reader will find more material in the further reading section, below. The laboratory evaluation of testicular function may be conveniently divided into assessment of spermatogenesis and assessment of endocrine gonadal function. Semen analysis is used both in the evaluation of male fertility and the follow-up of treatment regimens for male subfertility. It must be noted that there is wide inter-laboratory variation in the results, although this has been reduced in recent years by automated systems using electro-optics or computer-assisted analysis. Furthermore, there is marked variation in sperm output on a day-to-day basis. It is often more useful in clinical practice to assess the actual fertilization capacity of sperm in vitro. A semen sample is collected by masturbation and should ideally be analysed within 1 h. Basic semen analysis measures the number of spermatozoa (per unit volume and per ejaculate), motility and morphology. These are assessed according to World Health Organization (WHO) criteria, updated in 2010, although it should be noted that individual laboratories will also have developed their own reference ranges. The WHO reference ranges were determined by the analysis of semen from fertile men whose partners became pregnant within one year of trying to conceive. Average sperm count is > 60 million/mL and a count of < 15 million/mL is considered subfertile. There should be > 39 million sperm in a given ejaculate and semen volume should be 1.5 mL or greater. Motility is defined as the percentage of moving sperm and is subdivided into the following categories: • non-progressive motility (NP): all other patterns of motility with an absence of progression • immotility (IM): no movement. The WHO reference values state that ≥ 40% of the sperm should exhibit either PR or NP motility and ≥ 32% should exhibit PR motility. At least 58% of sperm should be alive – this is especially important for samples with a PR motility of < 40% and must be assessed within 1 h of ejaculation. Morphology, describing defects in the appearance of spermatozoa, is most commonly assessed according to the WHO morphology standards. The WHO lower reference limit for normal morphology is 4%. Few samples exhibit > 25% normal spermatozoa. Other parameters measured include volume, pH, white blood cell count, antisperm antibodies, colour, viscosity, agglutination, aggregation and time until liquefaction. Reference ranges for semen analysis are shown in Table 23.2. TABLE 23.2 Reference ranges for semen variablesa a WHO classification described in text. PR, progressive motility; NP, non-progressive motility. Determination of basal plasma LH and FSH concentrations is a routine part of the assessment of the hypothalamo–pituitary–gonadal axis. As LH and, to a lesser extent, FSH are secreted in a pulsatile fashion, more accurate interpretation may be possible if serial samples are taken over a period of time. However, in routine clinical practice, the measurement of basal plasma concentrations of LH and FSH with testosterone provides most of the information required. Elevated LH and FSH in the presence of low testosterone concentrations suggest primary testicular failure; low or normal LH and FSH associated with low testosterone concentrations suggest a hypothalamo–pituitary disorder. It is important to recognize that the latter may be caused by any acute illness. Rarely, LH alone may be elevated with a low testosterone (the Sertoli-only syndrome) and, in patients with abnormal spermatogenesis, FSH may be raised with a normal testosterone. It should be remembered that pituitary and gonadal dysfunction may coexist, as in haemochromatosis. In this situation, LH, FSH and testosterone concentrations are low. The GnRH test (Appendix 23.1i) has been used to assess pituitary reserve of LH and FSH and overall hypothalamo–pituitary function. This test is now used relatively infrequently, as it rarely provides extra information beyond that from basal LH and FSH concentrations. The clomifene stimulation test (Appendix 23.1ii) can be used to assess the hypothalamo–pituitary axis with respect to gonadotrophin secretion. Clomifene competitively inhibits hypothalamo–pituitary androgen binding. The normal response is a doubling of LH and FSH by day 10 after administration. A low response, however, does not distinguish between abnormalities at the pituitary or hypothalamic level. Plasma testosterone concentrations exhibit diurnal variation throughout the day – some studies suggesting a variation of as much as 30% between a sample taken at 08.00 h and one at 16.00 h. Consequently, and by convention, testosterone is usually determined on a sample taken before 09.00 h. Analysis of serum testosterone concentrations must take into account sex hormone-binding globulin (SHBG), as 50% of circulating testosterone is bound to SHBG. Low testosterone concentrations are seen both in primary testicular disease, indicating poor Leydig cell function, and hypothalamo–pituitary disorders with depressed gonadotrophin secretion. Elevated concentrations are seen in patients with gonadotrophin- or androgen-secreting tumours (e.g. Leydig cell tumours), in whom gonadotrophin concentrations will be low. The hCG stimulation test (Appendix 23.1
Reproductive function in the male
INTRODUCTION
THE TESTES
Production and actions of testosterone
Hypothalamo–pituitary control of testicular function
Testicular malignancy
Endocrine disrupting chemicals
EVALUATION OF TESTICULAR FUNCTION
Semen analysis
Variable
Reference range
Volume
> 1.5 mL
pH
> 7.2
Sperm concentration
> 15 × 106 sperm/mL
Total count
> 39 × 106 sperm/ejaculate
Motility
40% or more with total motility (PR + NP)
32% or more with PR motility
Vitality
58% or more alive
White blood cells
< 1 × 106 cells/ejaculate
Fructose
> 13 μmol/ejaculate
Endocrine evaluation: hypothalamo–pituitary–gonadal axis
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


