EYES: THE PHOTORECEPTOR SYSTEM
Accessory Structures of the Eye
EARS: THE VESTIBULOAUDITORY SYSTEM
Information about the external world is conveyed to the central nervous system (CNS) from sensory receptors. Chemoreceptive cells for the senses of taste and smell were discussed with the digestive and respiratory system (see Chapters 15 and 17, respectively) and the various mechanoreceptors that mediate the sense of touch were presented with the skin (see Chapter 18). This chapter describes the eye, both its photoreceptors and auxiliary structures, and the ear which mediates the senses of equilibrium and hearing via mechanoreceptors in the vestibulocochlear apparatus.
EYES: THE PHOTORECEPTOR SYSTEM
Eyes (Figure 23–1) are highly developed photosensitive organs for analyzing the form, intensity, and color of light reflected from objects and providing the sense of sight. Protected within the orbits of the skull which also contain adipose cushions, each eyeball consists externally of a tough, fibrous globe that maintains its overall shape. Internally the eye contains transparent tissues that refract light to focus the image, a layer of photosensitive cells, and a system of neurons that collect, process, and transmit visual information to the brain.
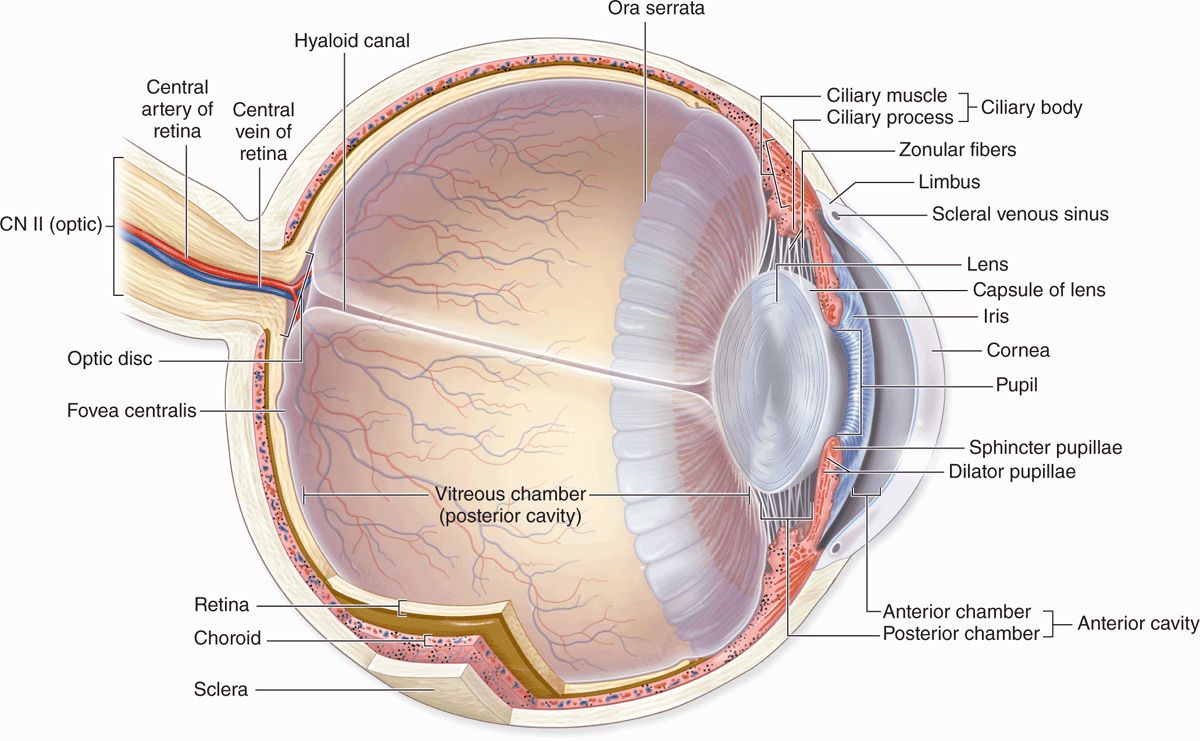
FIGURE 23–1 Internal anatomy of the eye.
Each eye is composed of three concentric tunics or layers (Table 23–1):
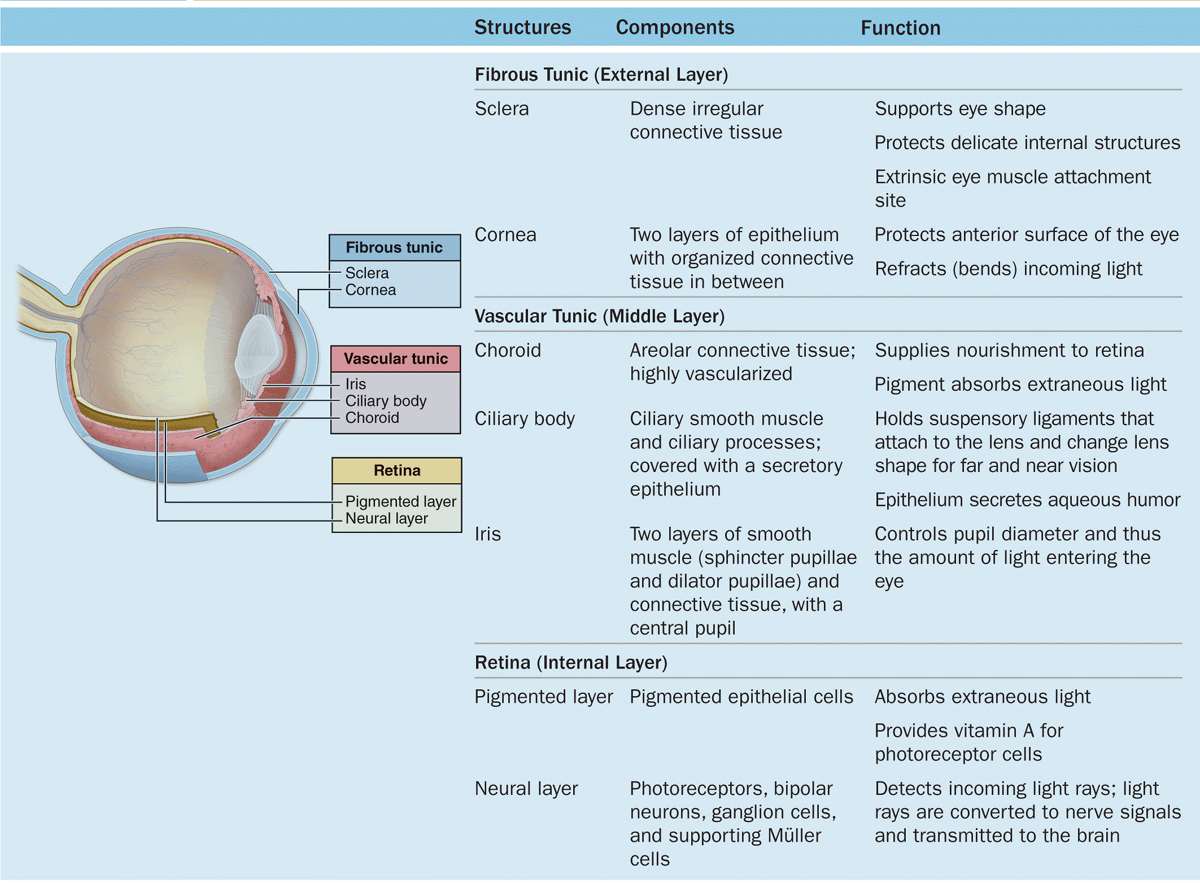
 A tough external fibrous layer consisting of the sclera and the transparent cornea;
A tough external fibrous layer consisting of the sclera and the transparent cornea;
 A middle vascular layer that includes the choroid, ciliary body, and iris; and
A middle vascular layer that includes the choroid, ciliary body, and iris; and
 An inner sensory layer, the retina, which communicates with the cerebrum through the posterior optic nerve (Figure 23–1).
An inner sensory layer, the retina, which communicates with the cerebrum through the posterior optic nerve (Figure 23–1).
Not part of these layers, the lens is a perfectly transparent biconvex structure held in place by a circular system of zonular fibers that attach it to the ciliary body and by close apposition to the posterior vitreous body (Figure 23–1). Partly covering the anterior surface of the lens is an opaque pigmented extension of the middle layer called the iris, which surrounds a central opening, the pupil (Figure 23–1).
Located in the anterior portion of the eye, the iris and lens are bathed in clear aqueous humor that fills both the anterior chamber between the cornea and iris and the posterior chamber between the iris and lens (Figure 23–1). Aqueous humor flows through the pupil that connects these two chambers.
The posterior vitreous chamber, surrounded by the retina, lies behind the lens and its zonular fibers and contains a large gelatinous mass of transparent connective tissue called the vitreous body.
Important aspects of embryonic eye formation are shown in Figure 23–2 and include the following:
 In the 4-week embryo epithelial optic vesicles bulge bilaterally from the forebrain, then elongate as the optic stalks bearing optic cups (Figure 23–2a).
In the 4-week embryo epithelial optic vesicles bulge bilaterally from the forebrain, then elongate as the optic stalks bearing optic cups (Figure 23–2a).
 Inductive interactions between the optic cups and the overlying surface ectoderm cause the latter to invaginate and eventually detach as the initially hollow lens vesicles (Figure 23–2b).
Inductive interactions between the optic cups and the overlying surface ectoderm cause the latter to invaginate and eventually detach as the initially hollow lens vesicles (Figure 23–2b).
 The optic stalk develops as the optic nerve and in an inferior groove called the choroid fissure encloses the hyaloid vessels that supply blood for the developing lens and optic cup (Figure 23–2c).
The optic stalk develops as the optic nerve and in an inferior groove called the choroid fissure encloses the hyaloid vessels that supply blood for the developing lens and optic cup (Figure 23–2c).
 In the ensuing weeks, head mesenchyme differentiates to form most of the tissue in the eye’s two outer layers and the vitreous. Ectoderm of the optic cup differentiates as the retina and surface ectoderm forms the corneal epithelium (Figure 23–2d). When the lens is fully formed, the distal hyaloid artery and vein disappear, leaving only the blood supply to the retina.
In the ensuing weeks, head mesenchyme differentiates to form most of the tissue in the eye’s two outer layers and the vitreous. Ectoderm of the optic cup differentiates as the retina and surface ectoderm forms the corneal epithelium (Figure 23–2d). When the lens is fully formed, the distal hyaloid artery and vein disappear, leaving only the blood supply to the retina.
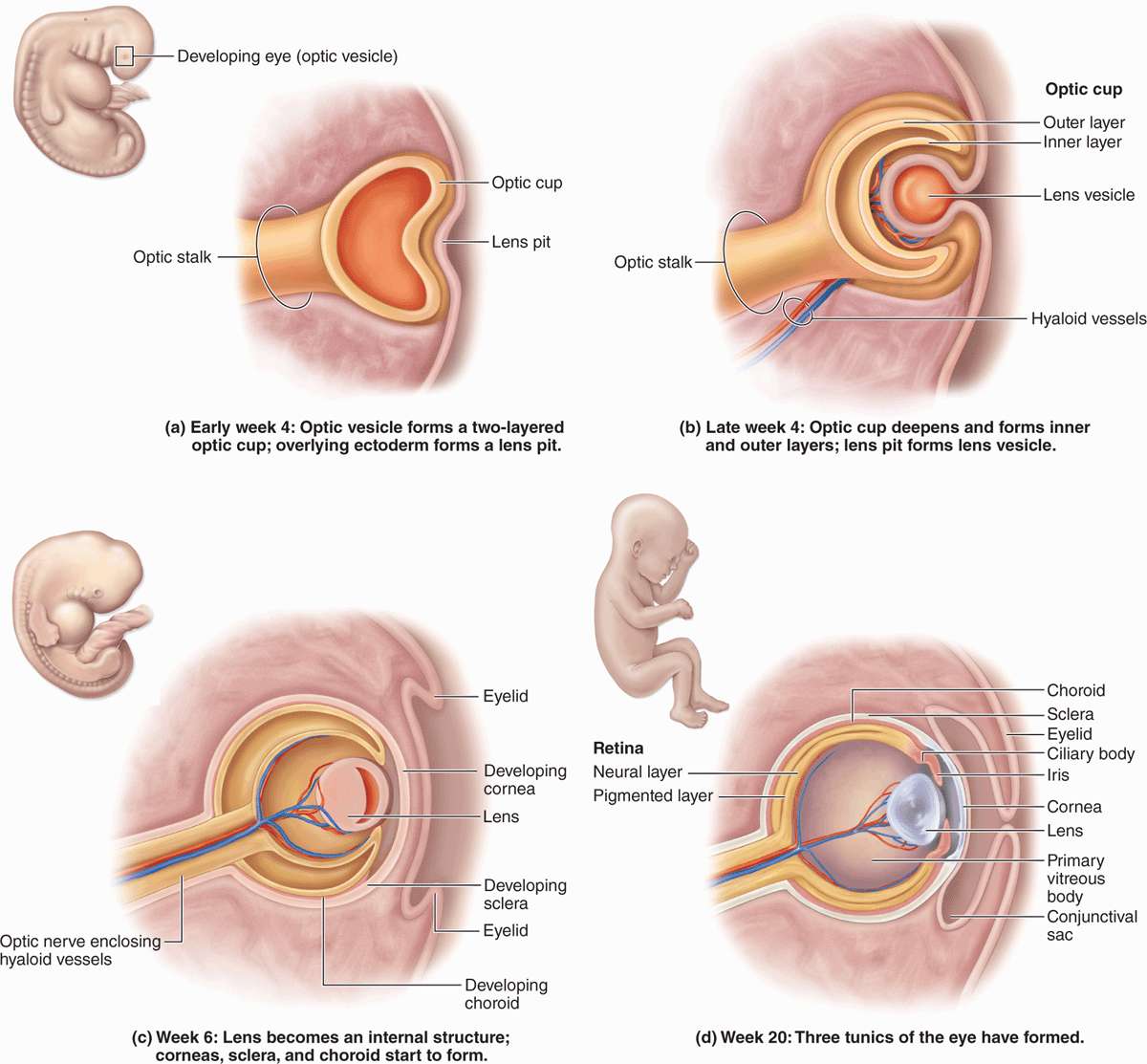
FIGURE 23–2 Development of eye.
Fibrous Layer
This layer includes two major regions, the posterior sclera and anterior cornea, joined at the limbus.
Sclera
The fibrous, external layer of the eyeball protects the more delicate internal structures and provides sites for muscle insertion (Table 23–1). The white posterior five-sixths of this layer is the sclera (Figure 23–1), which encloses a portion of the eyeball about 22 mm in diameter in adults. The sclera averages 0.5 mm in thickness and consists mainly of dense connective tissue, with flat bundles of type I collagen parallel to the organ surface but intersecting in various directions; microvasculature is present near the outer surface.
Tendons of the extraocular muscles which move the eyes insert into the anterior region of the sclera. Posteriorly the sclera thickens to approximately 1 mm and joins with the epineurium covering the optic nerve. Where it surrounds the choroid, the sclera includes an inner suprachoroid lamina, with less collagen, more fibroblasts, elastic fibers, and melanocytes.
Cornea
In contrast to the sclera, the anterior one-sixth of the eye—the cornea—is transparent and completely avascular (Figure 23–1). A section of the cornea shows five distinct layers:
 An external stratified squamous epithelium;
An external stratified squamous epithelium;
 An anterior limiting membrane (Bowman’s membrane), which is the basement membrane of the external stratified epithelium;
An anterior limiting membrane (Bowman’s membrane), which is the basement membrane of the external stratified epithelium;
 The thick stroma;
The thick stroma;
 A posterior limiting membrane (Descemet’s membrane), which is the basement membrane of the endothelium; and
A posterior limiting membrane (Descemet’s membrane), which is the basement membrane of the endothelium; and
 An inner simple squamous endothelium.
An inner simple squamous endothelium.
The stratified surface epithelium is nonkeratinized, five or six cell layers thick, and comprises about 10% of the corneal thickness (Figure 23–3). The basal cells have a high proliferative capacity important for renewal and repair of the corneal surface and emerge from stem cells in the corneoscleral limbus that encircles the cornea. The flattened surface cells have microvilli protruding into a protective tear film of lipid, glycoprotein, and water. As another protective adaptation, the corneal epithelium also has one of the richest sensory nerve supplies of any tissue.
The basement membrane of this epithelium, often called Bowman’s membrane, is very thick (8-10 μm) and contributes to the stability and strength of the cornea, helping to protect against infection of the underlying stroma.
The stroma, or substantia propria, makes up 90% of the cornea’s thickness and consists of approximately 60 layers of parallel collagen bundles aligned at approximately right angles to each other and extending almost the full diameter of the cornea. The uniform orthogonal array of collagen fibrils contributes to the transparency of this avascular tissue. Between the collagen lamellae are cytoplasmic extensions of flattened fibroblast-like cells called keratocytes (Figure 23–3). The ground substance around these cells contains proteoglycans such as lumican, with keratan sulfate and chondroitin sulfate, which help maintain the precise organization and spacing of the collagen fibrils.
 MEDICAL APPLICATION
MEDICAL APPLICATION
The shape or curvature of the cornea can be changed surgically to improve certain visual abnormalities involving the ability to focus. In the common ophthalmologic procedure, laser-assisted in situ keratomileusis (LASIK) surgery, the corneal epithelium is displaced as a flap and the stroma reshaped by an excimer laser which vaporizes collagen and keratocytes in a highly controlled manner with no damage to adjacent cells or ECM. After reshaping the stroma, the epithelial flap is repositioned and a relatively rapid regenerative response reestablishes normal corneal physiology. LASIK surgery is used to correct myopia (near-sightedness), hyperopia (far-sightedness), or astigmatism (irregular curvature of the cornea). Corneal grafts (transplants) between unrelated individuals can usually be accomplished successfully without immune rejection due in part to this tissue’s lack of both a vascular supply and lymphatic drainage and to local immune tolerance produced by ocular antigen-presenting cells and immunomodulatory factors in aqueous humor.
The posterior surface of the stroma is bounded by another thick basement membrane, called Descemet’s membrane, which supports the internal simple squamous corneal endothelium (Figure 23–3).
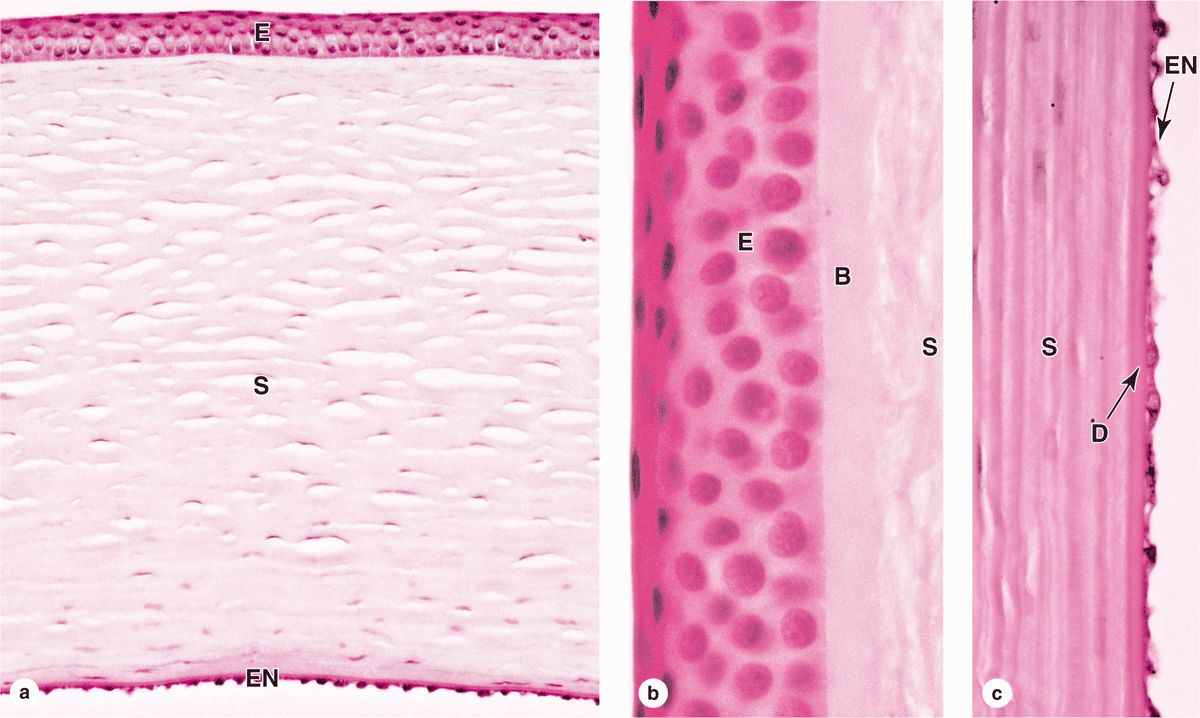
FIGURE 23–3 Cornea.
This endothelium maintains Descemet’s membrane and includes the most metabolically active cells of the cornea. Na+/K+ ATPase pumps in the basolateral membranes of these cells are largely responsible for regulating the proper hydration state of the corneal stroma to provide maximal transparency and optimal light refraction.
Limbus
Encircling the cornea is the limbus, a transitional area where the transparent cornea merges with the opaque sclera (Figures 23–1 and 23–4). Here Bowman’s membrane ends and the surface epithelium becomes more stratified as the conjunctiva that covers the anterior part of the sclera (and lines the eyelids). As mentioned previously, epithelial stem cells located at the limbus surface give rise to rapidly dividing progenitor cells that move centripetally into the corneal epithelium. The stroma becomes vascular and less well-organized at the limbus, as the collagen bundles merge with those of the sclera.
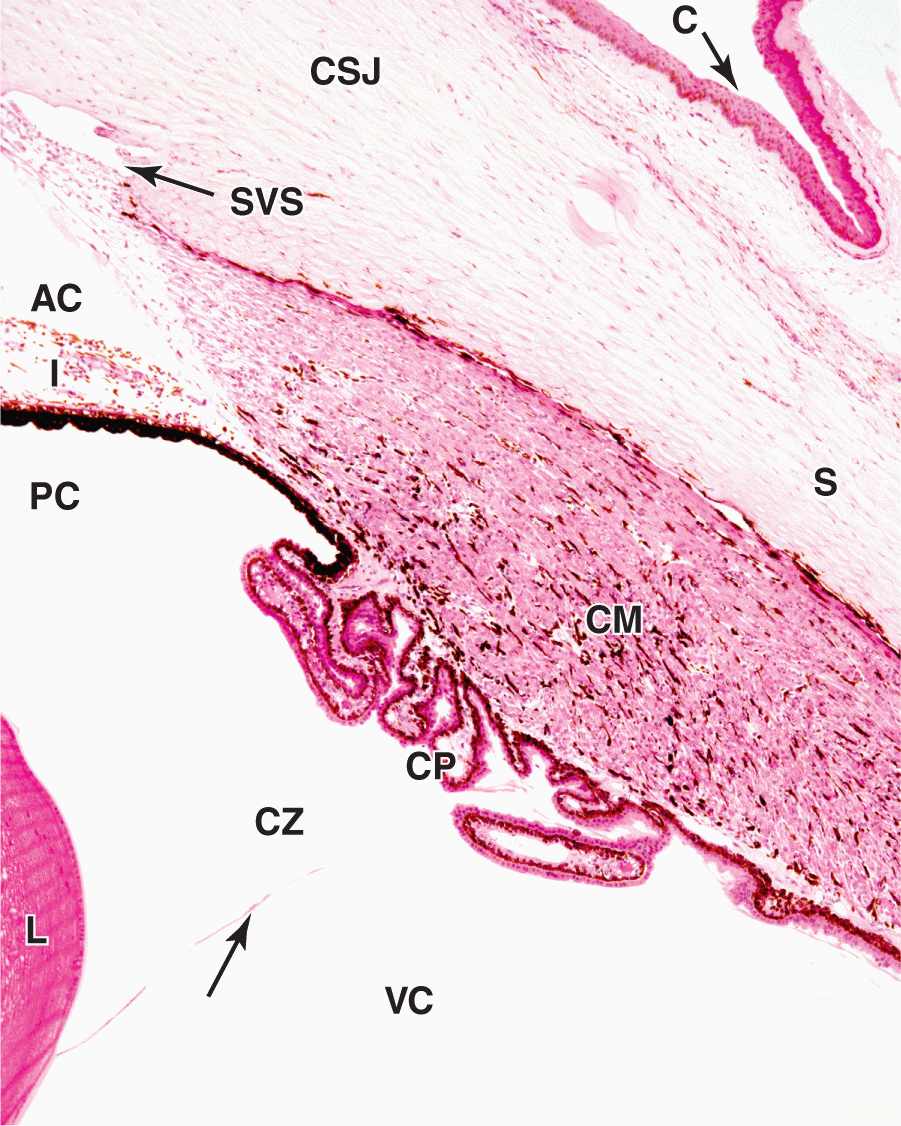
FIGURE 23–4 Corneoscleral junction (limbus) and ciliary body.
Also at the limbus Descemet’s membrane and its simple endothelium are replaced with a system of irregular endothelium-lined channels called the trabecular meshwork (Figure 23–5). These penetrate the stroma at the corneoscleral junction and allow slow, continuous drainage of aqueous humor from the anterior chamber. This fluid moves from these channels into the adjacent larger space of the scleral venous sinus, or canal of Schlemm (Figures 23–1, 23–4, and 23–5), which encircles the eye. From this sinus aqueous humor drains into small blood vessels (veins) of the sclera.
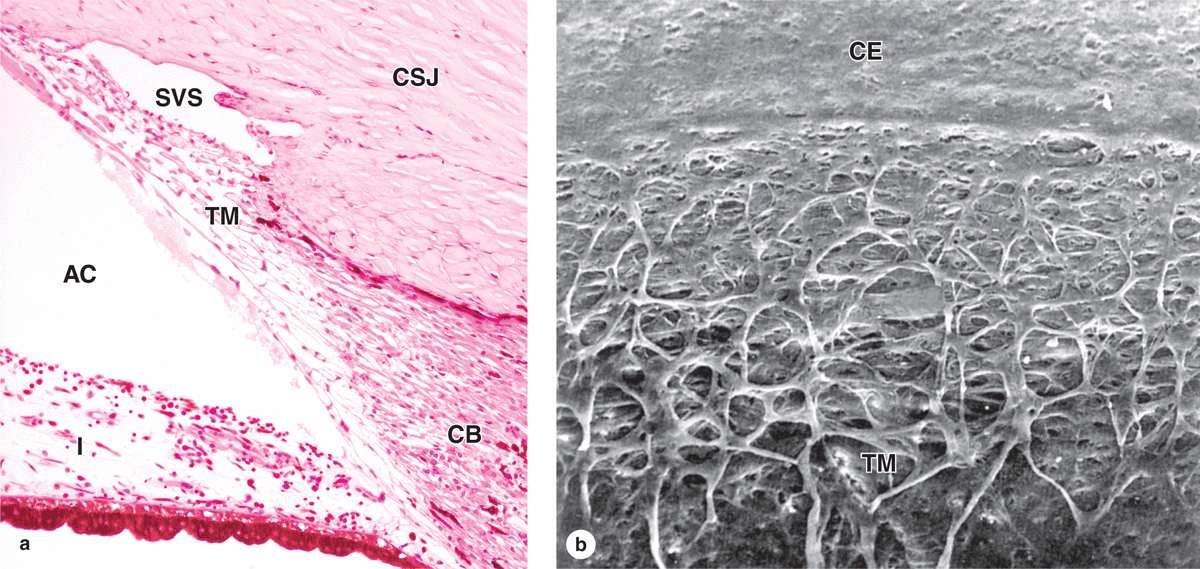
FIGURE 23–5 Trabecular meshwork and scleral venous sinus.
Vascular Layer
The eye’s more vascular middle layer, known as the uvea, consists of three parts, from posterior to anterior: the choroid, the ciliary body, and the iris (Table 23–1).
Choroid
Located in the posterior two-thirds of the eye, the choroid consists of loose, well-vascularized connective tissue and contains numerous melanocytes (Figure 23–6). These form a characteristic black layer in the choroid and prevent light from entering the eye except through the pupil. Two layers make up the choroid (Figure 23–6):
 The inner choroido-capillary lamina has a rich microvasculature important for nutrition of the outer retinal layers.
The inner choroido-capillary lamina has a rich microvasculature important for nutrition of the outer retinal layers.
 Bruch’s membrane, a thin extracellular sheet, is composed of collagen and elastic fibers surrounding the adjacent microvasculature and basal lamina of the retina’s pigmented layer.
Bruch’s membrane, a thin extracellular sheet, is composed of collagen and elastic fibers surrounding the adjacent microvasculature and basal lamina of the retina’s pigmented layer.
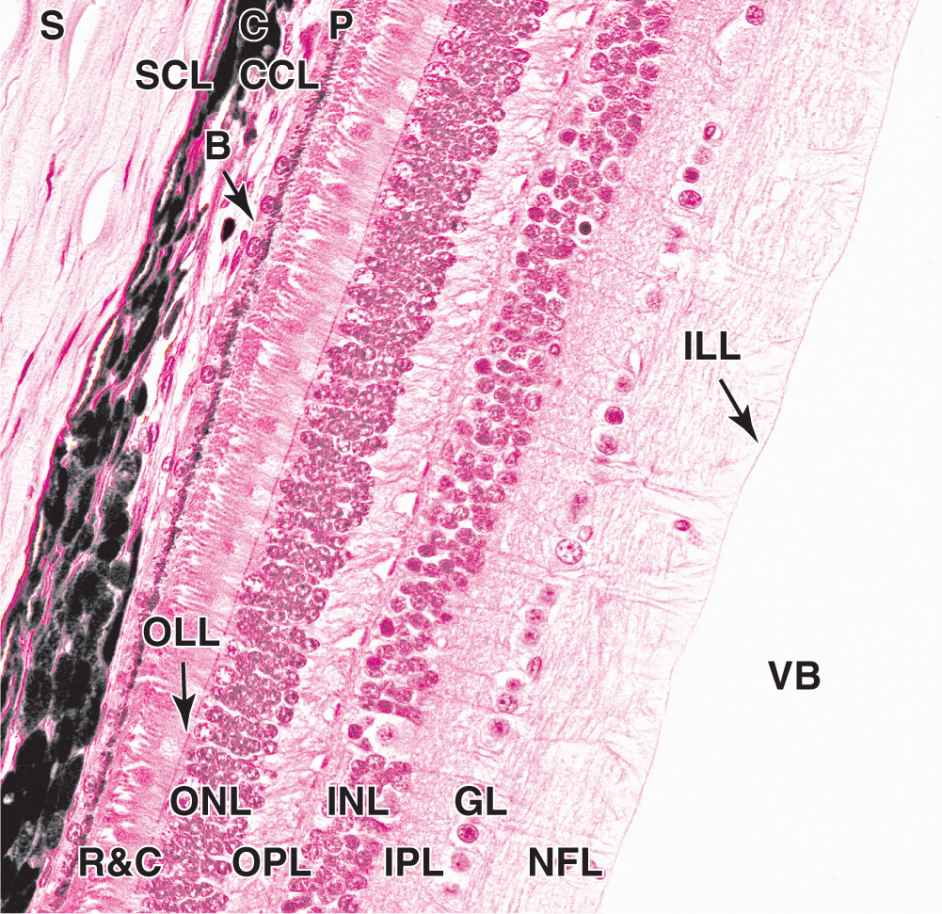
FIGURE 23–6 Sclera, choroid, and retina.
Ciliary Body
The ciliary body, the anterior expansion of the uvea that encircles the lens, lies posterior to the limbus (Figures 23–1 and 23–4). Like the choroid, most of the ciliary body rests on the sclera. Important structures associated with the ciliary body include the following:
 Ciliary muscle makes up most of the ciliary body’s stroma and consists of three groups of smooth muscle fibers. Contraction of these muscles affects the shape of the lens and is important in visual accommodation (see Lens).
Ciliary muscle makes up most of the ciliary body’s stroma and consists of three groups of smooth muscle fibers. Contraction of these muscles affects the shape of the lens and is important in visual accommodation (see Lens).
 Ciliary processes are a radially arranged series of about 75 ridges extending from the inner highly vascular region of the ciliary body. These provide a large surface area covered by a double layer of low columnar epithelial cells, the ciliary epithelium (Figure 23–7). The epithelial cells directly covering the stroma contain much melanin and correspond to the anterior projection of the pigmented retina epithelium. The surface layer of cells lacks melanin and is contiguous with the sensory layer of the retina.
Ciliary processes are a radially arranged series of about 75 ridges extending from the inner highly vascular region of the ciliary body. These provide a large surface area covered by a double layer of low columnar epithelial cells, the ciliary epithelium (Figure 23–7). The epithelial cells directly covering the stroma contain much melanin and correspond to the anterior projection of the pigmented retina epithelium. The surface layer of cells lacks melanin and is contiguous with the sensory layer of the retina.
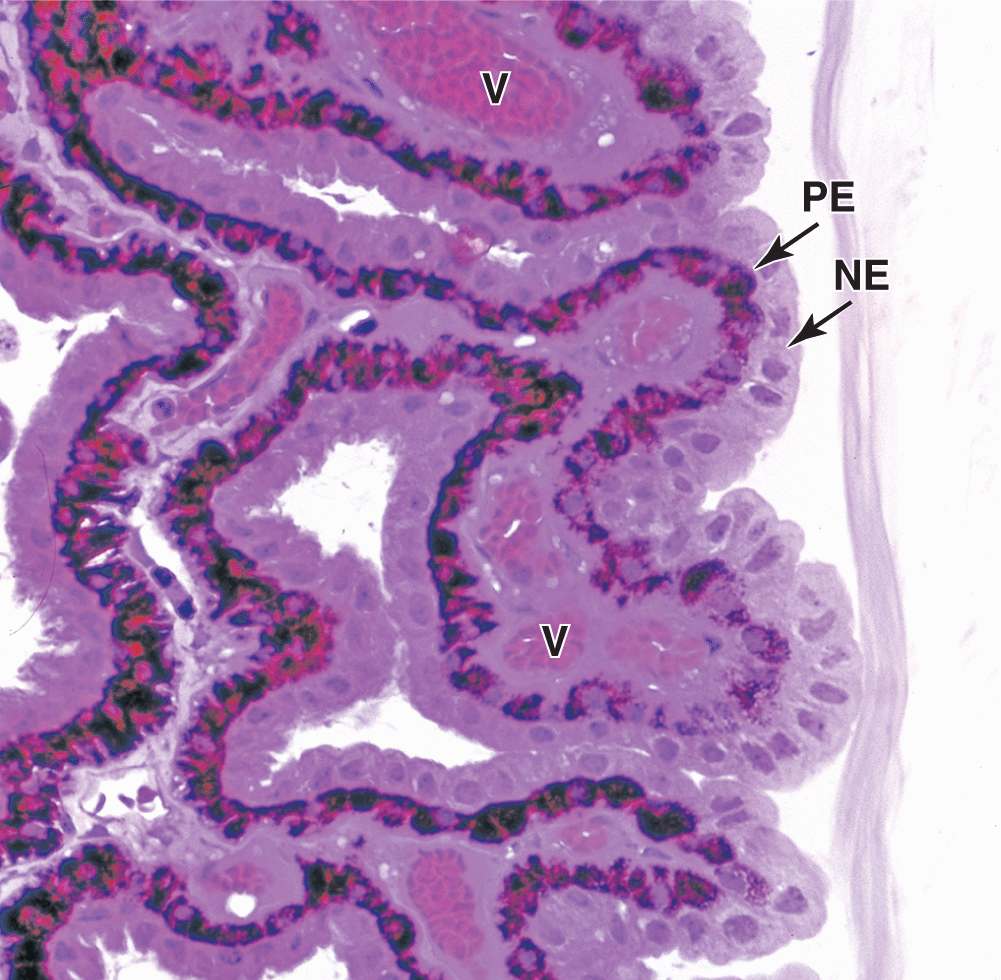
FIGURE 23–7 Epithelium of ciliary processes.
Cells of this dual epithelium have extensive basolateral folds with Na+/K+-ATPase activity and are specialized for secretion of aqueous humor. Fluid from the stromal microvasculature moves across this epithelium as aqueous humor, with an inorganic ion composition similar to that of plasma but almost no protein. As shown in Figure 23–8, aqueous humor is secreted by ciliary processes into the posterior chamber, flows through the pupil into the anterior chamber, and drains at the angle formed by the cornea and the iris into the channels of the trabecular meshwork and the scleral venous sinus, from which it enters venules of the sclera.
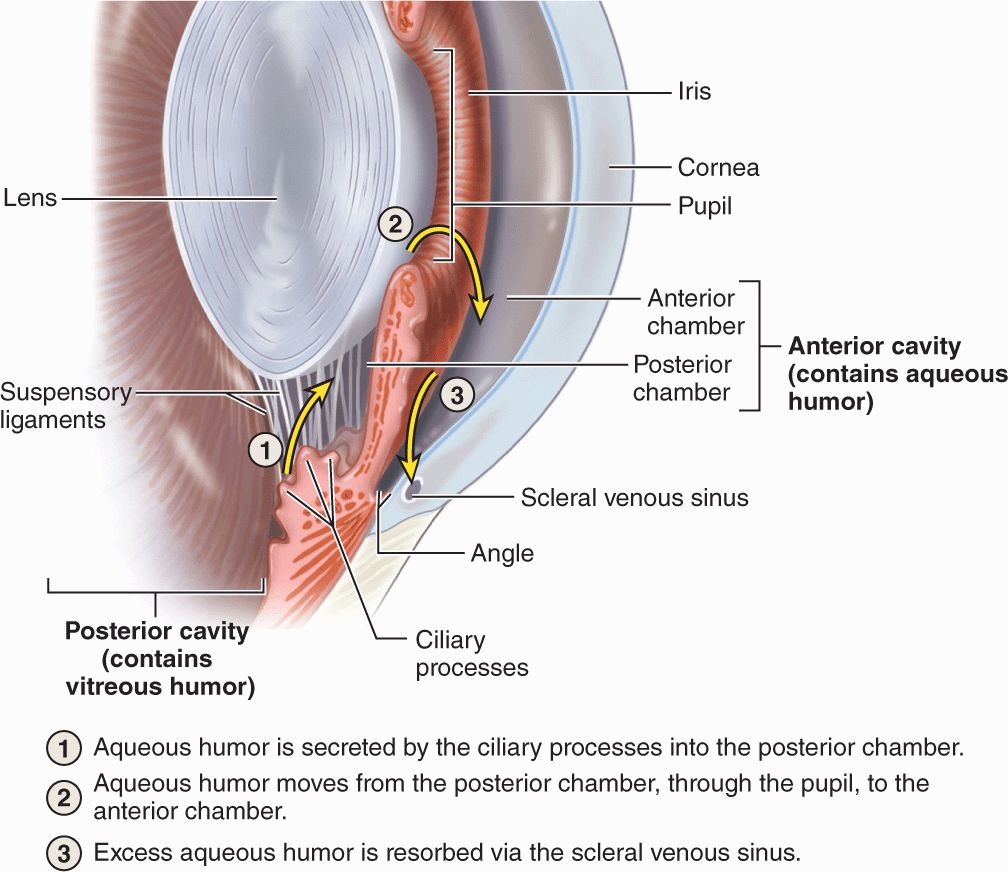
FIGURE 23–8 Production and removal of aqueous humor.
 The ciliary zonule is a system of many radially oriented fibers composed largely of fibrillin-1 and 2 produced by the nonpigmented epithelial cells on the ciliary processes. The fibers extend from grooves between the ciliary processes and attach to the surface of the lens (Figure 23–9), holding that structure in place.
The ciliary zonule is a system of many radially oriented fibers composed largely of fibrillin-1 and 2 produced by the nonpigmented epithelial cells on the ciliary processes. The fibers extend from grooves between the ciliary processes and attach to the surface of the lens (Figure 23–9), holding that structure in place.
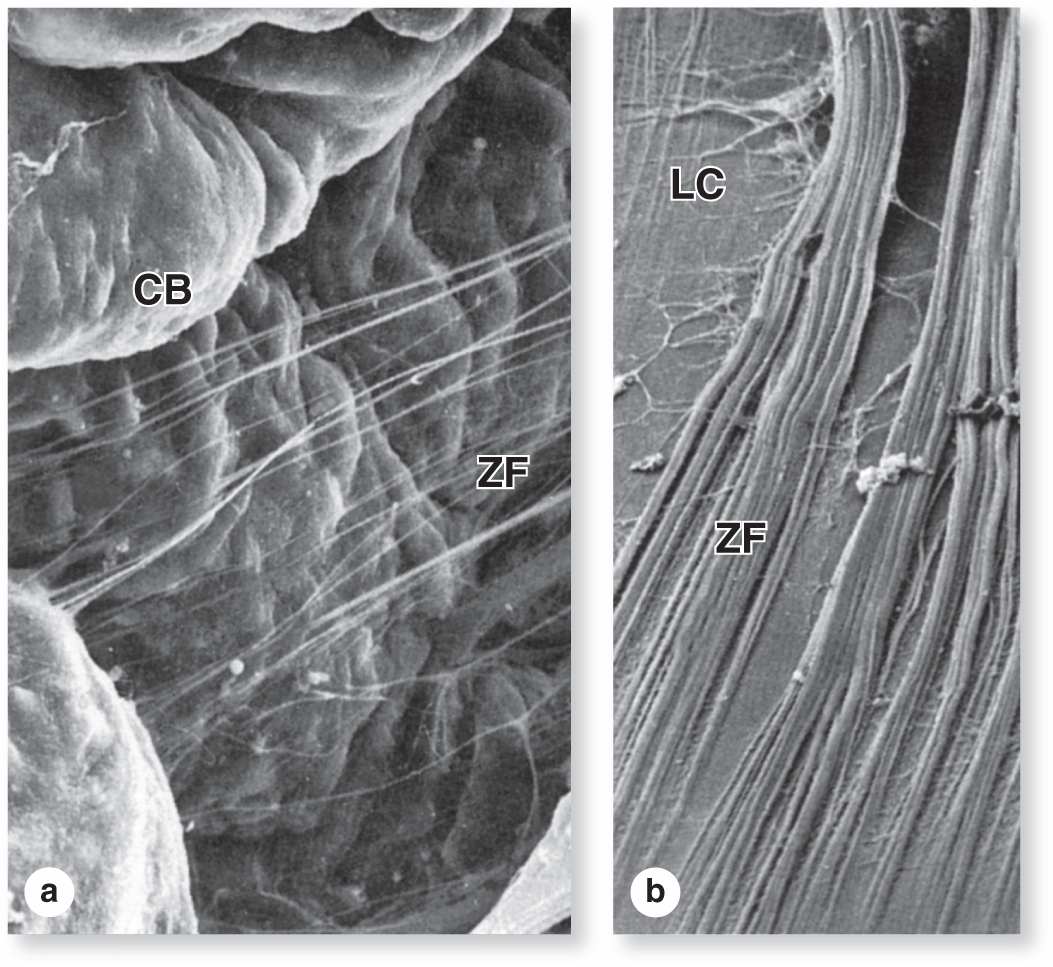
FIGURE 23–9 Ciliary zonule fibers.
 MEDICAL APPLICATION
MEDICAL APPLICATION
Aqueous humor is produced continuously. If its drainage from the anterior chamber is impeded, typically by obstruction of the trabecular meshwork or scleral venous sinus, intraocular pressure can increase, causing the condition called glaucoma. Untreated glaucoma can cause pressing of the vitreous body against the retina, affecting visual function and possibly leading to neuropathy in that tissue.
When the iridocorneal angle is more narrow than usual, the thickening of the peripheral iris that occurs with dilation of the pupil can occlude the angle and obstruct drainage of aqueous humor at the trabecular meshwork. This can result in the rapid development of intraocular hypertension known as angle closure glaucoma, acute glaucoma, or closed (narrow) angle glaucoma. This condition usually affects both eyes and causes blurred vision, eye pain, and headache. Treatment of this type of glaucoma usually includes some form of surgical intervention.
Iris
The iris is the most anterior extension of the middle uveal layer which covers part of the lens, leaving a round central pupil (Figure 23–1). The anterior surface of the iris, exposed to aqueous humor in the anterior chamber, consists of a dense layer of fibroblasts and melanocytes with interdigitating processes and is unusual for its lack of an epithelial covering (Figure 23–10a,b). Deeper in the iris, the stroma consists of loose connective tissue with melanocytes and sparse microvasculature.
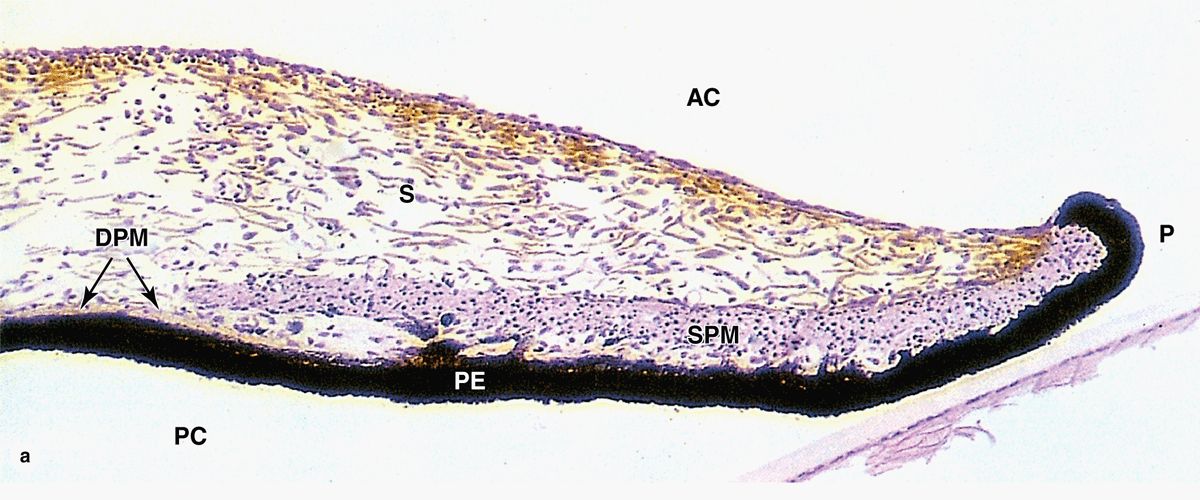
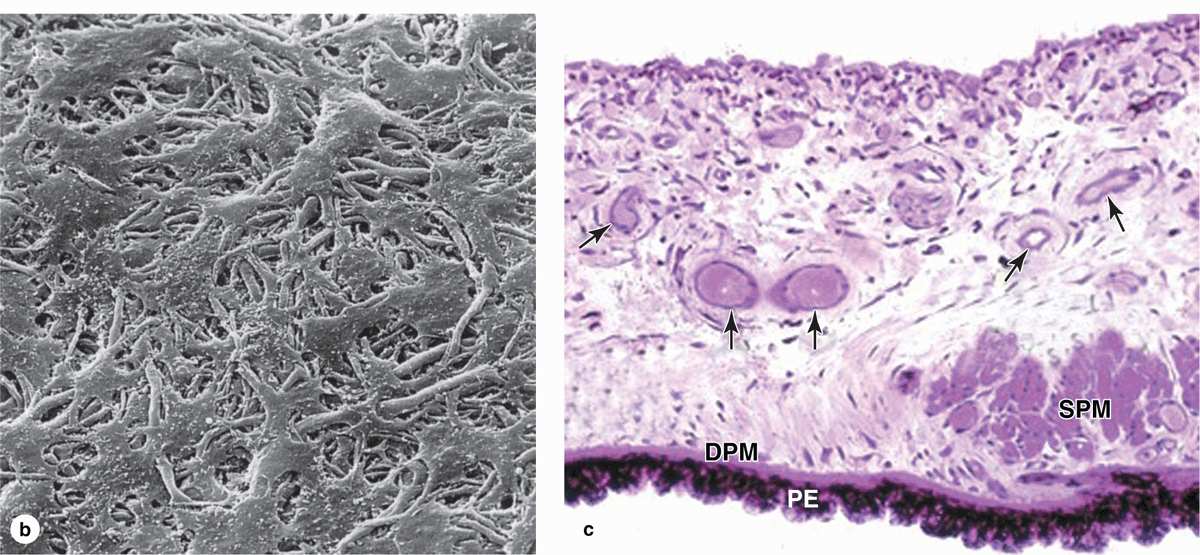
FIGURE 23–10 Iris.
The posterior surface of the iris has a two-layered epithelium continuous with that covering the ciliary processes, but very heavily filled with melanin. The highly pigmented posterior epithelium of the iris blocks all light from entering the eye except that passing through the pupil. Myoepithelial cells form a partially pigmented epithelial layer and extend contractile processes radially as the very thin dilator pupillae muscle (Figure 23–10). Smooth muscle fibers form a circular bundle near the pupil as the sphincter pupillae muscle. The dilator and sphincter muscles of the iris have sympathetic and parasympathetic innervation, respectively, for enlarging and constricting the pupil.
Melanocytes of the iris stroma provide the color of one’s eyes. In individuals with very few lightly pigmented cells in the stroma, light with a blue color is reflected back from the black pigmented epithelium on the posterior iris surface. As the number of melanocytes and density of melanin increase in the stroma, the iris color changes through various shades of green, gray, and brown. Individuals with albinism have almost no pigment and the pink color of their irises is due to the reflection of incident light from the blood vessels of the stroma.
Lens
The lens is a transparent biconvex structure suspended immediately behind the iris, which focuses light on the retina (see Figure 23–1). Derived from an invagination of the embryonic surface ectoderm (see Figure 23–2), the lens is a unique avascular tissue and is highly elastic, a property that normally decreases with age. The lens has three principal components:
 A thick (10-20 μm), homogeneous lens capsule composed of proteoglycans and type IV collagen surrounds the lens (Figure 23–11) and provides the place of attachment for the fibers of the ciliary zonule (Figure 23–10). This layer originates as the basement membrane of the embryonic lens vesicle.
A thick (10-20 μm), homogeneous lens capsule composed of proteoglycans and type IV collagen surrounds the lens (Figure 23–11) and provides the place of attachment for the fibers of the ciliary zonule (Figure 23–10). This layer originates as the basement membrane of the embryonic lens vesicle.
 A subcapsular lens epithelium consists of a single layer of cuboidal cells present only on the anterior surface of the lens (Figure 23–11). The epithelial cells attach basally to the surrounding lens capsule and their apical surfaces bind to the internal lens fibers. At the posterior edge of this epithelium, near the equator of the lens, the epithelial cells divide to provide new cells that differentiate as lens fibers. This process allows for growth of the lens and continues at a slow, decreasing rate near the equator of the lens throughout adult life.
A subcapsular lens epithelium consists of a single layer of cuboidal cells present only on the anterior surface of the lens (Figure 23–11). The epithelial cells attach basally to the surrounding lens capsule and their apical surfaces bind to the internal lens fibers. At the posterior edge of this epithelium, near the equator of the lens, the epithelial cells divide to provide new cells that differentiate as lens fibers. This process allows for growth of the lens and continues at a slow, decreasing rate near the equator of the lens throughout adult life.
 Lens fibers are highly elongated, terminally differentiated cells that appear as thin, flattened structures (Figure 23–11). Developing from cells in the lens epithelium, lens fibers typically become 7 to 10 mm long, with cross-section dimensions of only 2 by 8 μm. The cytoplasm becomes filled with a group of proteins called crystallins, and the organelles and nuclei undergo autophagy. Lens fibers are packed tightly together and form a perfectly transparent tissue highly specialized for light refraction.
Lens fibers are highly elongated, terminally differentiated cells that appear as thin, flattened structures (Figure 23–11). Developing from cells in the lens epithelium, lens fibers typically become 7 to 10 mm long, with cross-section dimensions of only 2 by 8 μm. The cytoplasm becomes filled with a group of proteins called crystallins, and the organelles and nuclei undergo autophagy. Lens fibers are packed tightly together and form a perfectly transparent tissue highly specialized for light refraction.
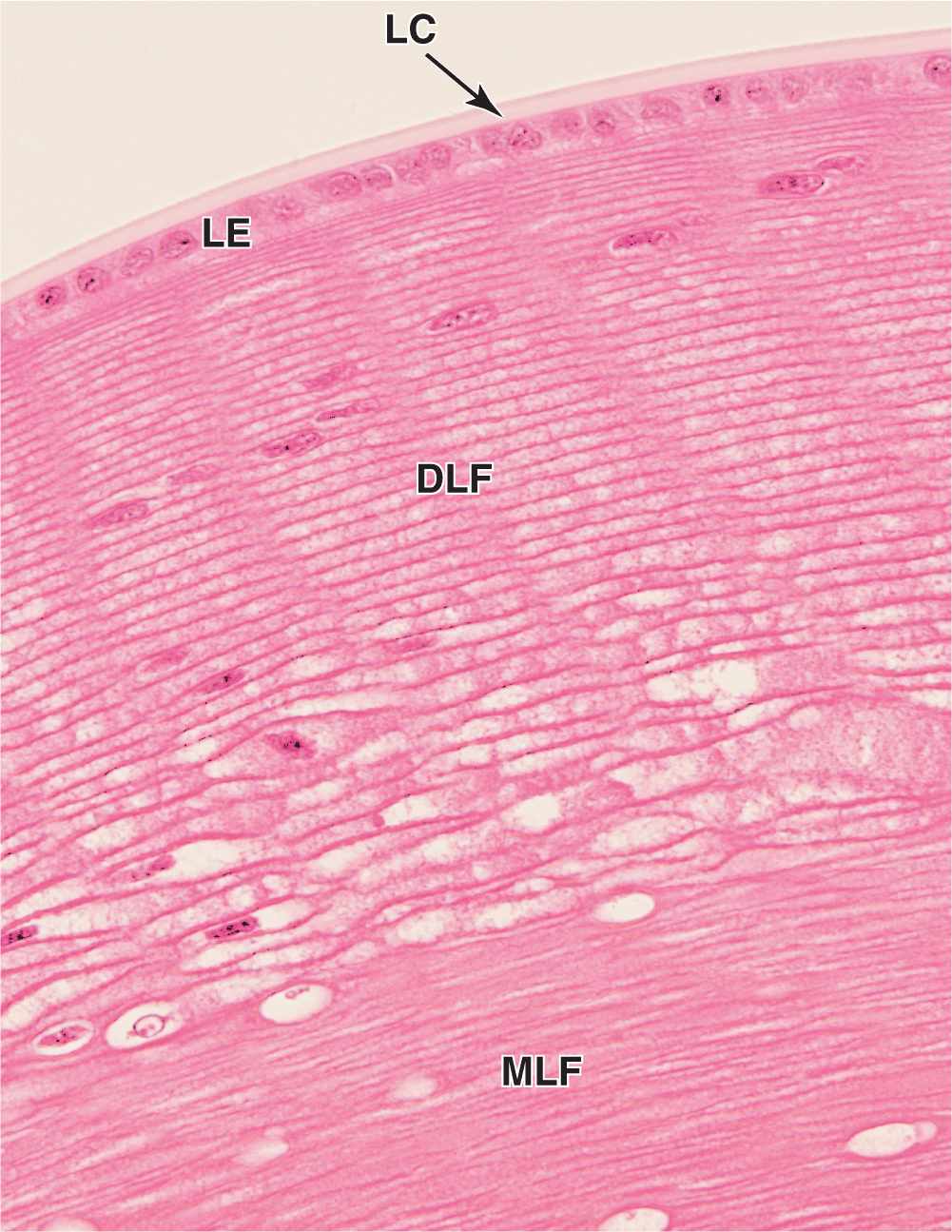
FIGURE 23–11 Lens.
The lens is held in place by fibers of the ciliary zonule, which extend from the lens capsule to the ciliary body (Figures 23–1 and 23–9). Together with the ciliary muscles, this structure allows the process of visual accommodation, which permits focusing on near and far objects by changing the curvature of the lens (Figure 23–12). When the eye is at rest or gazing at distant objects, ciliary muscles relax and the resulting shape of the ciliary body puts tension on the zonule fibers, which pulls the lens into a flatter shape. To focus on a close object the ciliary muscles contract, causing forward displacement of the ciliary body, which relieves some of the tension on the zonule and allows the lens to return to a more rounded shape and keep the object in focus. In the fourth decade of life presbyopia (Gr. presbyter, elder + L. opticus, relating to eyes) normally causes the lenses to lose elasticity and their ability to undergo accommodation.
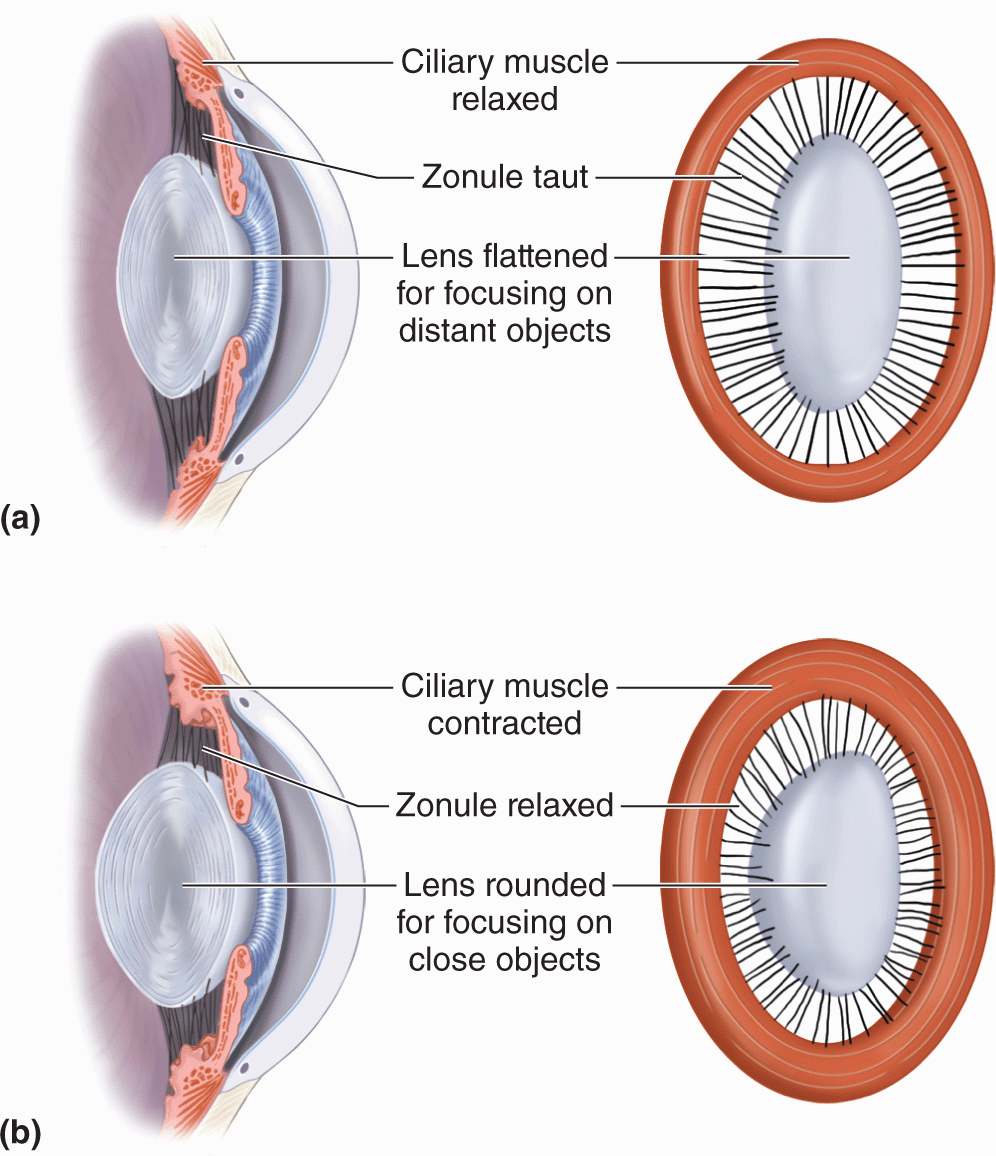
FIGURE 23–12 Accommodation of the lens.
 MEDICAL APPLICATION
MEDICAL APPLICATION
Presbyopia is corrected by wearing glasses with convex lenses (reading glasses). In older individuals, denaturation of crystallins commonly begins to occur in lens fibers, making them less transparent. When areas of the lens become opaque or cloudy and vision is impaired, the condition is termed a cataract. Causes of cataract include excessive exposure to ultraviolet light or other radiation, trauma, and as secondary effects in diseases such as diabetes mellitus and hypertension.
In modern cataract surgery the lens is removed by aspiration of the lens substance while it is emulsified by a vibrating probe. The posterior side of the lens capsule and its inserted zonular fibers are left in place at the posterior chamber. The concave posterior capsule, or capsular bag, is then used as the site for implantation of an acrylic intraocular lens (IOL) prosthesis. Research is under way to develop IOLs capable of natural accommodation.
Vitreous Body
The vitreous body occupies the large vitreous chamber behind the lens (see Figure 23–1). It consists of transparent, gel-like connective tissue that is 99% water (vitreous humor), with collagen fibrils and hyaluronate, contained within an external lamina called the vitreous membrane. The only cells in the vitreous body are a small mesenchymal population near the membrane called hyalocytes, which synthesize the hyaluronate and collagen, and a few macrophages.
Retina
The retina, the innermost tunic of the eye, develops with two fundamental sublayers from the inner and outer layers of embryonic optic cup (see Figure 23–2 and Table 23–1):
 The outer pigmented layer is a simple cuboidal epithelium attached to Bruch’s membrane and the choroidocapillary lamina of the choroid (see Figure 23–6). This heavily pigmented layer forms the other part of the dual epithelium covering the ciliary body and posterior iris.
The outer pigmented layer is a simple cuboidal epithelium attached to Bruch’s membrane and the choroidocapillary lamina of the choroid (see Figure 23–6). This heavily pigmented layer forms the other part of the dual epithelium covering the ciliary body and posterior iris.
 The inner retinal region, the neural layer, is thick and stratified with various neurons and photoreceptors. Although its neural structure and visual function extend anterior only as far as the ora serrata (see Figure 23–1), this layer continues as part of the dual cuboidal epithelium that covers the surface of the ciliary body and posterior iris.
The inner retinal region, the neural layer, is thick and stratified with various neurons and photoreceptors. Although its neural structure and visual function extend anterior only as far as the ora serrata (see Figure 23–1), this layer continues as part of the dual cuboidal epithelium that covers the surface of the ciliary body and posterior iris.
 MEDICAL APPLICATION
MEDICAL APPLICATIONThe pigmented epithelium and the photoreceptor layer of the retina, derived from the two layers of the optic cup, are not firmly joined to each other. Head trauma or other conditions can cause the two layers to separate with an intervening space. In such regions of detached retina, the photoreceptor cells no longer have access to metabolic support from the pigmented layer and choroid and will eventually die. Prompt repositioning of the retina and reattaching it with laser surgery is an effective treatment.
Retina Pigmented Epithelium
The pigmented epithelial layer consists of cuboidal or low columnar cells with basal nuclei and surrounds the neural layer of the retina. The cells have well-developed junctional complexes, gap junctions, and numerous invaginations of the basal membranes associated with mitochondria. The apical ends of the cells extend processes and sheath-like projections that surround the tips of the photoreceptors. Melanin granules are numerous in these extensions and in the apical cytoplasm (Figure 23–13). This cellular region also contains numerous phagocytic vacuoles and secondary lysosomes, peroxisomes, and abundant smooth ER (SER) specialized for retinal (vitamin A) isomerization. The diverse functions of the retinal pigmented epithelium include the following:
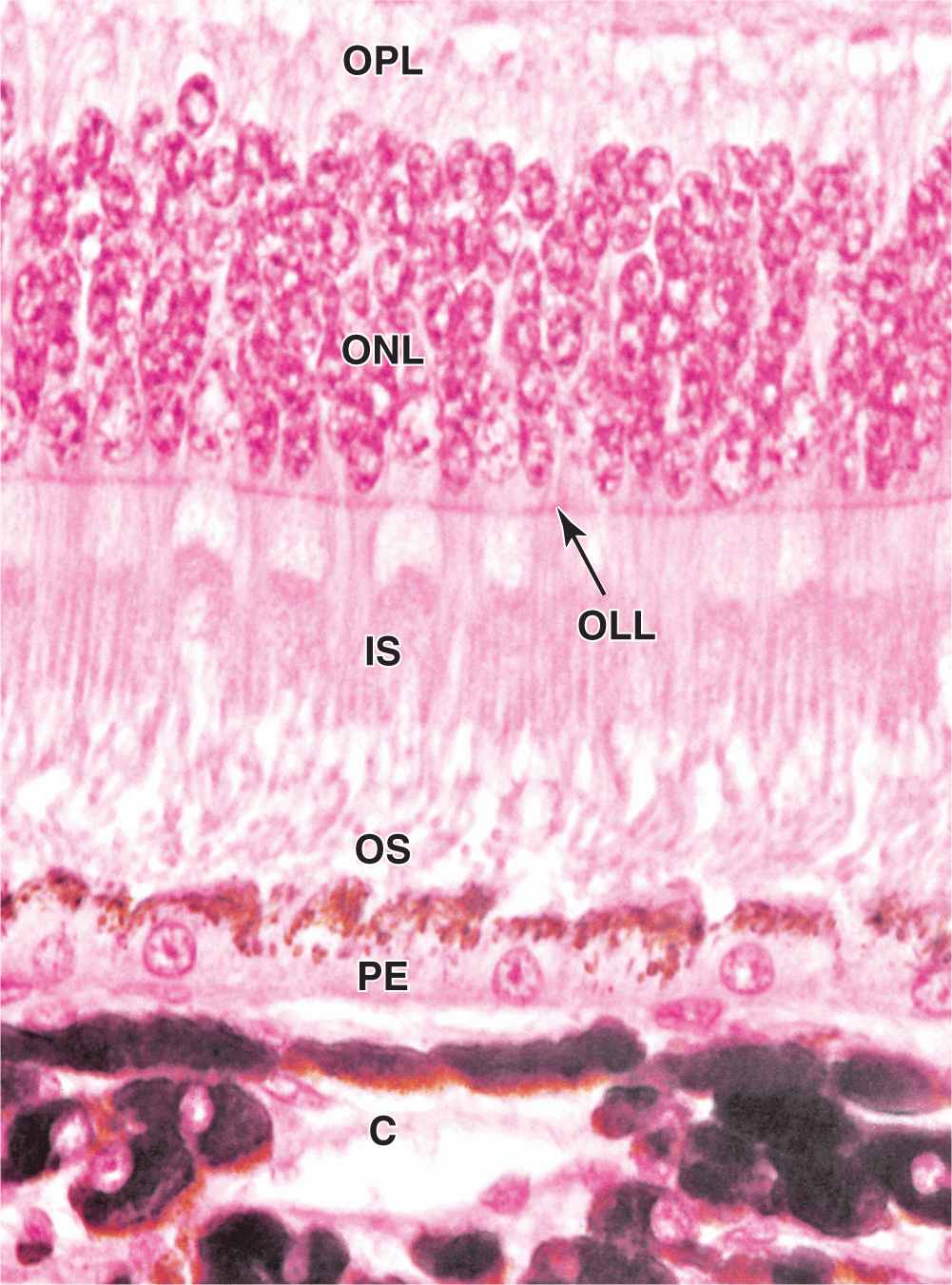
FIGURE 23–13 Pigmented epithelium of retina.



