GENERAL STRUCTURE OF THE DIGESTIVE TRACT
The digestive system consists of the digestive tract— oral cavity, esophagus, stomach, small and large intestines, and anus—and its associated glands—salivary glands, liver, and pancreas (Figure 15–1). Also called the gastrointestinal (Gi) tract or alimentary canal, its function is to obtain from ingested food the molecules necessary for the maintenance, growth, and energy needs of the body. During digestion proteins, complex carbohydrates, nucleic acids, and fats are broken down into their small molecule subunits that are easily absorbed through the small intestine lining. Most water and electrolytes are absorbed in the large intestine. In addition, the inner layer of the entire digestive tract forms an important protective barrier between the content of the tract’s lumen and the internal milieu of the body’s connective tissue and vasculature.
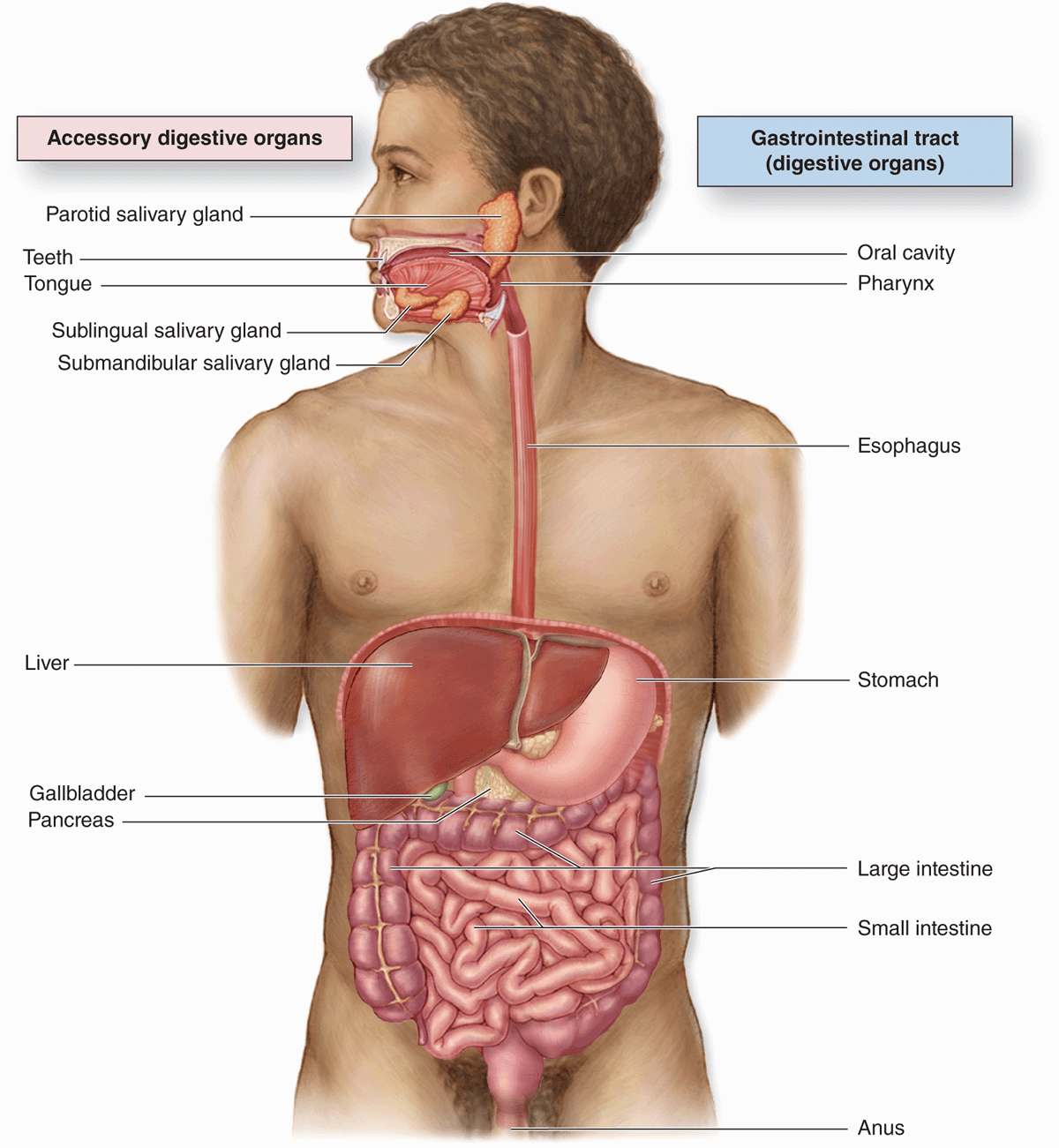
FIGURE 15–1 The digestive system.
Structures within the digestive tract allow the following:
 ingestion, or introduction of food and liquid into the oral cavity.
ingestion, or introduction of food and liquid into the oral cavity.
 Mastication, or chewing, which divides solid food into digestible pieces.
Mastication, or chewing, which divides solid food into digestible pieces.
 Motility, muscular movements of materials through the tract.
Motility, muscular movements of materials through the tract.
 Secretion of lubricating and protective mucus, digestive enzymes, acidic and alkaline fluids, and bile.
Secretion of lubricating and protective mucus, digestive enzymes, acidic and alkaline fluids, and bile.
 Hormone release for local control of motility and secretion.
Hormone release for local control of motility and secretion.
 Chemical digestion or enzymatic degradation of large macromolecules in food to smaller molecules and their subunits.
Chemical digestion or enzymatic degradation of large macromolecules in food to smaller molecules and their subunits.
 Absorption of the small molecules and water into the blood and lymph.
Absorption of the small molecules and water into the blood and lymph.
 Elimination of indigestible, unabsorbed components of food.
Elimination of indigestible, unabsorbed components of food.
GENERAL STRUCTURE OF THE DIGESTIVE TRACT
All regions of the GI tract have certain structural features in common. The GI tract is a hollow tube with a lumen of variable diameter and a wall made up of four main layers: the mucosa, submucosa, muscularis, and serosa. Figure 15–2 shows a general overview of these four layers; key features of each layer are summarized here.
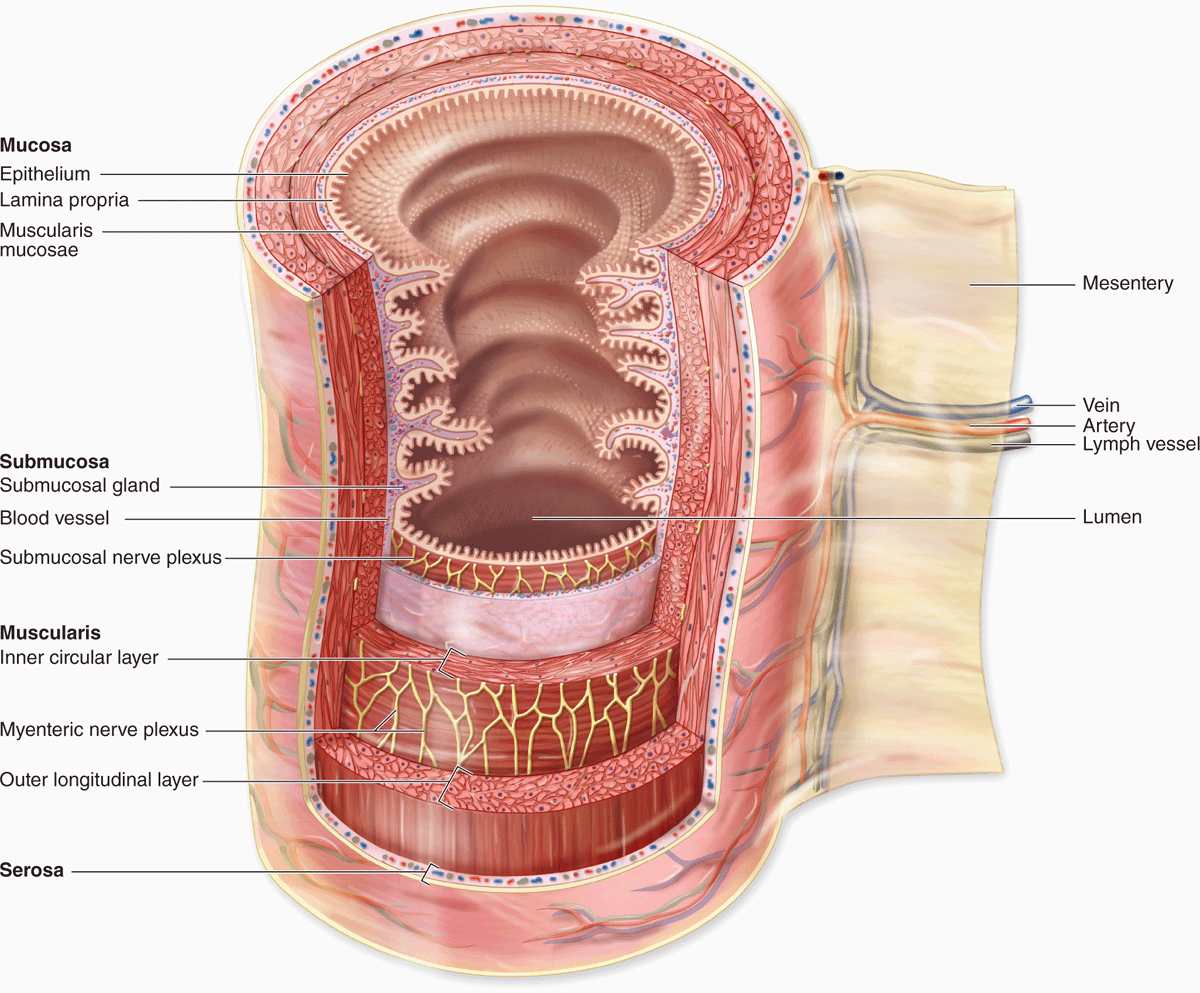
FIGURE 15–2 Major layers and organization of the digestive tract.
 The mucosa consists of an epithelial lining; an underlying lamina propria of loose connective tissue rich in blood vessels, lymphatics, lymphocytes, smooth muscle cells, and often containing small glands; and a thin layer of smooth muscle called the muscularis mucosae separating mucosa from submucosa and allowing local movements of the mucosa. The mucosa is also frequently called a mucous membrane.
The mucosa consists of an epithelial lining; an underlying lamina propria of loose connective tissue rich in blood vessels, lymphatics, lymphocytes, smooth muscle cells, and often containing small glands; and a thin layer of smooth muscle called the muscularis mucosae separating mucosa from submucosa and allowing local movements of the mucosa. The mucosa is also frequently called a mucous membrane.
 The submucosa contains denser connective tissue with larger blood and lymph vessels and the submucosal (Meissner) plexus of autonomic nerves. It may also contain glands and significant lymphoid tissue.
The submucosa contains denser connective tissue with larger blood and lymph vessels and the submucosal (Meissner) plexus of autonomic nerves. It may also contain glands and significant lymphoid tissue.
 The thick muscularis (or muscularis externa) is composed of smooth muscle cells organized as two or more sublayers. In the internal sublayer (closer to the lumen), the fiber orientation is generally circular; in the external sublayer it is longitudinal. The connective tissue between the muscle sublayers contains blood and lymph vessels, as well as the myenteric (Auerbach) nerve plexus of many autonomic neurons aggregated into small ganglia and interconnected by pre- and postganglionic nerve fibers. This and the submucosal plexus together comprise the enteric nervous system of the digestive tract. Contractions of the muscularis, which mix and propel the luminal contents forward, are generated and coordinated by the myenteric plexus.
The thick muscularis (or muscularis externa) is composed of smooth muscle cells organized as two or more sublayers. In the internal sublayer (closer to the lumen), the fiber orientation is generally circular; in the external sublayer it is longitudinal. The connective tissue between the muscle sublayers contains blood and lymph vessels, as well as the myenteric (Auerbach) nerve plexus of many autonomic neurons aggregated into small ganglia and interconnected by pre- and postganglionic nerve fibers. This and the submucosal plexus together comprise the enteric nervous system of the digestive tract. Contractions of the muscularis, which mix and propel the luminal contents forward, are generated and coordinated by the myenteric plexus.
 The serosa is a thin layer of loose connective tissue, rich in blood vessels, lymphatics, and adipose tissue, with a simple squamous covering epithelium or mesothelium. In the abdominal cavity, the serosa is continuous with mesenteries, thin membranes covered by mesothelium on both sides that support the intestines. Mesenteries are continuous with the peritoneum, a serous membrane that lines that cavity. In places where the digestive tract is not suspended in a cavity but bound directly to adjacent structures, such as in the esophagus (Figure 15–1), the serosa is replaced by a thick adventitia, a connective tissue layer that merges with the surrounding tissues and lacks mesothelium.
The serosa is a thin layer of loose connective tissue, rich in blood vessels, lymphatics, and adipose tissue, with a simple squamous covering epithelium or mesothelium. In the abdominal cavity, the serosa is continuous with mesenteries, thin membranes covered by mesothelium on both sides that support the intestines. Mesenteries are continuous with the peritoneum, a serous membrane that lines that cavity. In places where the digestive tract is not suspended in a cavity but bound directly to adjacent structures, such as in the esophagus (Figure 15–1), the serosa is replaced by a thick adventitia, a connective tissue layer that merges with the surrounding tissues and lacks mesothelium.
The numerous free immune cells and lymphoid nodules in the mucosa and submucosa constitute the MALT described in Chapter 14. The digestive tract normally contains thousands of microbial species, including both useful inhabitants of the gut as well as potential pathogens ingested with food and drink. The mucosa-associated immune defense system provides an essential backup to the thin physical barrier of the epithelial lining. Located just below the epithelium, the lamina propria is rich with macrophages and lymphocytes, many for production of IgA antibodies. Such antibodies undergo transcytosis into the intestinal lumen bound to the secretory protein produced by the epithelial cells. This IgA complex resists proteolysis by digestive enzymes and provides important protection against specific viral and bacterial pathogens.
 MEDICAL APPLICATION
MEDICAL APPLICATION
In diseases such as Hirschsprung disease (congenital aganglionic megacolon) or Chagas disease (trypanosomiasis, infection with the protozoan Trypanosoma cruzi), plexuses in the digestive tract’s enteric nervous system are absent or severely injured, respectively. This disturbs digestive tract motility and produces dilations in some areas. The rich autonomic innervation of the enteric nervous system also provides an anatomic explanation of the well-known actions of emotional stress on the stomach and other regions of the GI tract.
ORAL CAVITY
The oral cavity (Figure 15–1) is lined with stratified squamous epithelium, which may be keratinized, partially keratinized, or nonkeratinized depending on the location. Epithelial differentiation, keratinization, and the interface between the epithelium and lamina propria are similar to those features in the epidermis and dermis and are discussed more extensively with skin (see Chapter 18). Like the keratinized surface cells of epidermis, the flattened superficial cells of the oral epithelium undergo continuous desquamation, or loss at the surface. Unlike those of the epidermis, the shed cells of the nonkeratinized or parakeratinized oral epithelium retain their nuclei.
 MEDICAL APPLICATION
MEDICAL APPLICATION
Viral infections with herpes simplex 1 cause death of infected epithelial cells that can lead to vesicular or ulcerating lesions of the oral mucosa or skin near the mouth. In the oral cavity such areas are called canker sores, and on the skin they are usually called cold sores or fever blisters. Such lesions, often painful and clustered, occur when the immune defenses are weakened by emotional stress, fever, illness, or local skin damage, allowing the virus, present in the local nerves, to move into the epithelial cells.
The keratinized cell layers resist damage from abrasion and are best developed in the masticatory mucosa on the gingiva (gum) and hard palate. The lamina propria in these regions rests directly on the periosteum of underlying bone. Nonkeratinized squamous epithelium predominates in the lining mucosa over the soft palate, cheeks, the floor of the mouth, and the pharynx, the posterior region of the oral cavity leading to the esophagus. Lining mucosa overlies a thick submucosa containing many minor salivary glands, which secrete continuously to keep the mucosal surface wet, and diffuse lymphoid tissue. Throughout the oral cavity, the epithelium contains transient antigen-presenting cells and rich sensory innervation.
The well-developed core of striated muscle in the lips, or labia, (Figure 15–3) makes these structures highly mobile for ingestion, speech, and other forms of communication. Both lips have three differently covered surfaces:
FIGURE 15–3 lip.
 The internal mucous surface has lining mucosa with a thick, nonkeratinized epithelium and many minor labial salivary glands.
The internal mucous surface has lining mucosa with a thick, nonkeratinized epithelium and many minor labial salivary glands.
 The red vermilion zone of each lip is covered by very thin keratinized stratified squamous epithelium and is transitional between the oral mucosa and skin. This region lacks salivary or sweat glands and is kept moist with saliva from the tongue. The underlying connective tissue is very rich in both sensory innervation and capillaries, which impart the pink color to this region.
The red vermilion zone of each lip is covered by very thin keratinized stratified squamous epithelium and is transitional between the oral mucosa and skin. This region lacks salivary or sweat glands and is kept moist with saliva from the tongue. The underlying connective tissue is very rich in both sensory innervation and capillaries, which impart the pink color to this region.
 The outer surface has thin skin, consisting of epidermal and dermal layers, sweat glands, and many hair follicles with sebaceous glands.
The outer surface has thin skin, consisting of epidermal and dermal layers, sweat glands, and many hair follicles with sebaceous glands.
Tongue
The tongue is a mass of striated muscle covered by mucosa, which manipulates ingested material during mastication and swallowing. The muscle fibers are oriented in all directions, allowing a high level of mobility. Connective tissue between the small fascicles of muscle is penetrated by the lamina propria, which makes the mucous membrane strongly adherent to the muscular core. The lower surface of the tongue is smooth, with typical lining mucosa. The dorsal surface is irregular, having hundreds of small protruding papillae of various types on its anterior two-thirds and the massed lingual tonsils on the posterior third, or root of the tongue (Figure 15–4). The papillary and tonsillar areas of the lingual surface are separated by a V-shaped groove called the sulcus terminalis.
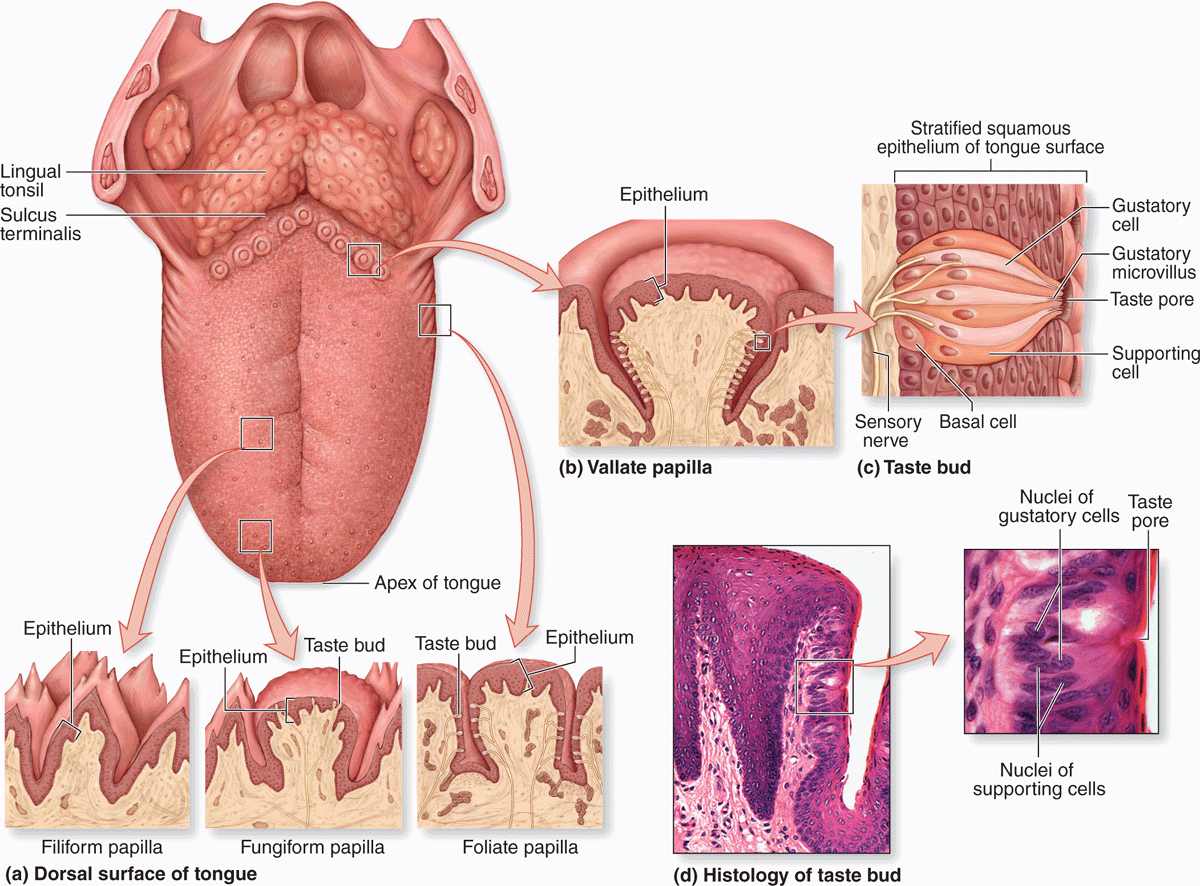
FIGURE 15–4 Tongue, lingual papillae, and taste buds.
The lingual papillae are elevations of the mucous membrane that assume various forms and functions. There are four types (Figure 15–4):
 Filiform papillae (Figure 15–5) are very numerous, have an elongated conical shape, and are heavily keratinized, which gives their surface a gray or whitish appearance. They provide a rough surface that facilitates movement of food during chewing.
Filiform papillae (Figure 15–5) are very numerous, have an elongated conical shape, and are heavily keratinized, which gives their surface a gray or whitish appearance. They provide a rough surface that facilitates movement of food during chewing.
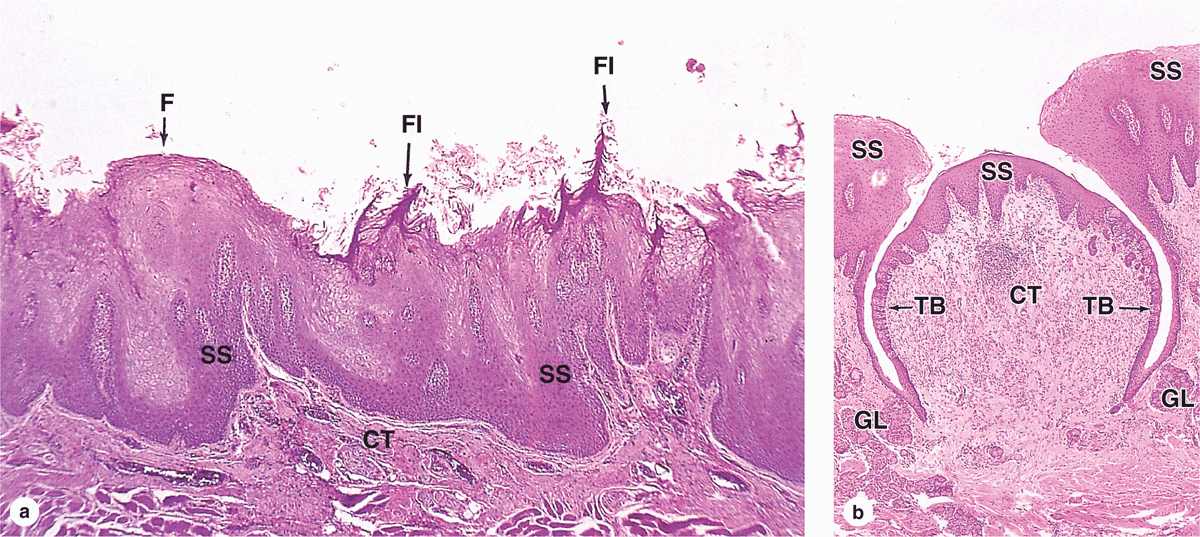
FIGURE 15–5 Lingual papillae.
 Fungiform papillae (Figure 15–5) are much less numerous, lightly keratinized, and interspersed among the filiform papillae. They are mushroom-shaped with well-vascularized and innervated cores of lamina propria.
Fungiform papillae (Figure 15–5) are much less numerous, lightly keratinized, and interspersed among the filiform papillae. They are mushroom-shaped with well-vascularized and innervated cores of lamina propria.
 Foliate papillae consist of several parallel ridges on each side of the tongue, anterior to the sulcus terminalis, but are rudimentary in humans, especially older individuals.
Foliate papillae consist of several parallel ridges on each side of the tongue, anterior to the sulcus terminalis, but are rudimentary in humans, especially older individuals.
 Vallate (or circumvallate) papillae (Figure 15–5) are the largest papillae, with diameters of 1 to 3 mm. Eight to twelve vallate papillae are normally aligned just in front of the terminal sulcus. Ducts of several small, serous salivary (von Ebner) glands empty into the deep, moatlike groove surrounding each vallate papilla. This provides a continuous flow of fluid over the taste buds that are abundant on the sides of these papillae, washing away food particles so that the taste buds can receive and process new gustatory stimuli. Secretions from these and other minor salivary glands associated with taste buds contain a lipase that prevents the formation of a hydrophobic film on these structures that would hinder gustation.
Vallate (or circumvallate) papillae (Figure 15–5) are the largest papillae, with diameters of 1 to 3 mm. Eight to twelve vallate papillae are normally aligned just in front of the terminal sulcus. Ducts of several small, serous salivary (von Ebner) glands empty into the deep, moatlike groove surrounding each vallate papilla. This provides a continuous flow of fluid over the taste buds that are abundant on the sides of these papillae, washing away food particles so that the taste buds can receive and process new gustatory stimuli. Secretions from these and other minor salivary glands associated with taste buds contain a lipase that prevents the formation of a hydrophobic film on these structures that would hinder gustation.
Taste buds are ovoid structures within the stratified epithelium on the tongue’s surface, which sample the general chemical composition of ingested material (Figures 15–4 and 15–5). Approximately 250 taste buds are present on the lateral surface of each vallate papilla, with many others present on fungiform and foliate (but not the keratinized filiform) papillae. They are not restricted to papillae and are also widely scattered elsewhere on the dorsal and lateral surfaces of the tongue, where they are also continuously flushed by numerous minor salivary glands.
A taste bud has 50 to 100 cells, about half of which are elongated gustatory (taste) cells, which turn over with a 7- to 10-day life span. Other cells present are slender supportive cells, immature cells, and slowly dividing basal stem cells that give rise to the other cell types. The base of each bud rests on the basal lamina and is entered by afferent sensory axons that form synapses with the gustatory cells. At the apical ends of the gustatory cells, microvilli project toward a 2-μm-wide opening in the structure called the taste pore. Molecules (tastants) dissolved in saliva contact the microvilli through the pore and interact with cell surface taste receptors (Figure 15–4).
Taste buds detect at least five broad categories of tastants: sodium ions (salty); hydrogen ions from acids (sour); sugars and related compounds (sweet); alkaloids and certain toxins (bitter); and amino acids such as glutamate and aspartate (umami; Jap. umami, savory). Salt and sour tastes are produced by ion channels and the other three taste categories are mediated by G-protein-coupled receptors. Receptor binding produces depolarization of the gustatory cells, stimulating the sensory nerve fibers that send information to the brain for processing. Conscious perception of tastes in food requires olfactory and other sensations in addition to taste bud activity.
Teeth
In the adult human there are normally 32 permanent teeth, arranged in two bilaterally symmetric arches in the maxillary and mandibular bones (Figure 15–6a). Each quadrant has eight teeth: two incisors, one canine, two premolars, and three permanent molars. Twenty of the permanent teeth are preceded by primary teeth (deciduous or milk teeth) that are shed; the others are permanent molars with no deciduous precursors. Each tooth has a crown exposed above the gingiva, a constricted neck at the gum, and one or more roots that fit firmly into bony sockets in the jaws called dental alveoli (Figure 15–6b).
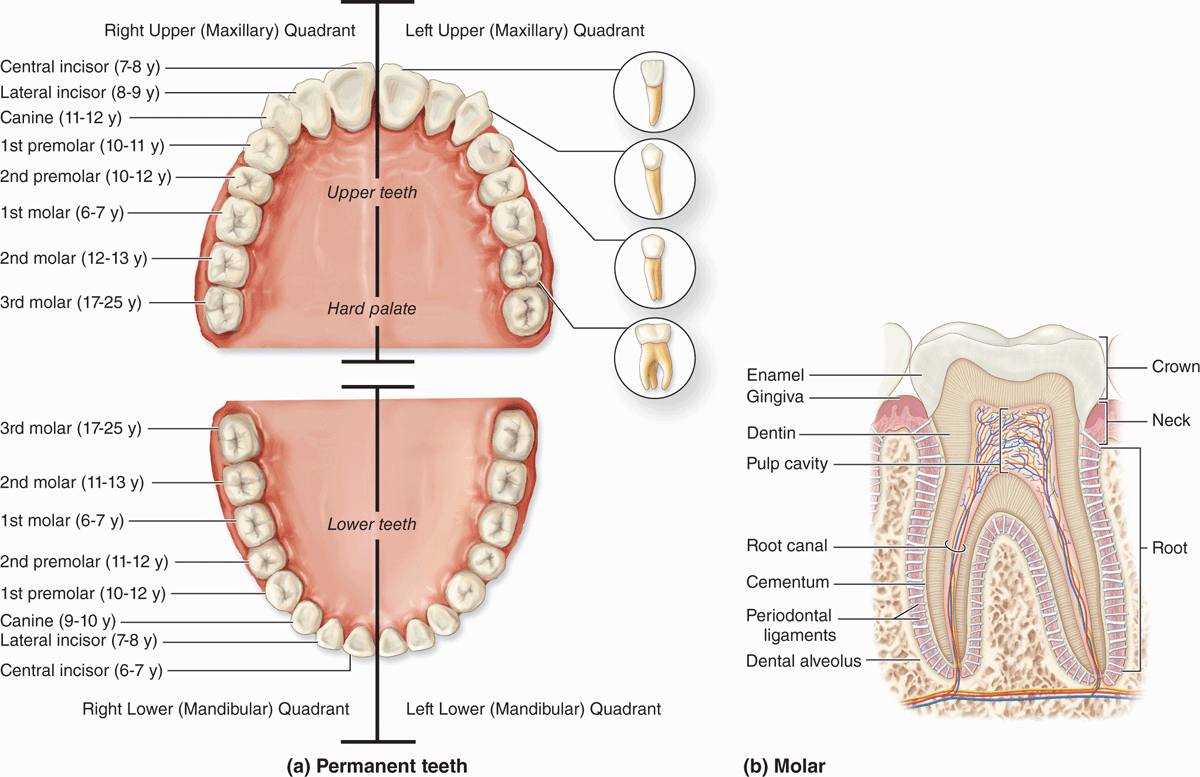
FIGURE 15–6 Teeth.
The crown is covered by very hard, acellular enamel and the roots by a bone-like tissue called cementum. These two coverings meet at the neck of the tooth. The bulk of a tooth is composed of another calcified material, dentin, which surrounds an internal pulp cavity (Figure 15–6b). Dental pulp is highly vascular and well-innervated and consists largely of loose, mesenchymal connective tissue with much ground substance, thin collagen fibers, fibroblasts, and mesenchymal stem cells. The pulp cavity narrows in each root as the root canal, which extends to an opening (apical foramen) at the tip of each root for the blood vessels, lymphatics, and nerves of the pulp cavity. The periodontal ligaments are fibrous connective tissue bundles of collagen fibers inserted into both the cementum and the alveolar bone.
Dentin
Dentin is a calcified tissue harder than bone, consisting of 70% hydroxyapatite. The organic matrix contains type I collagen and proteoglycans secreted from the apical ends of odontoblasts, tall polarized cells derived from the cranial neural crest that line the tooth’s pulp cavity (Figure 15-7a). Mineralization of the predentin matrix secreted by odontoblasts involves matrix vesicles in a process similar to that occurring in osteoid during bone formation (see Chapter 8).
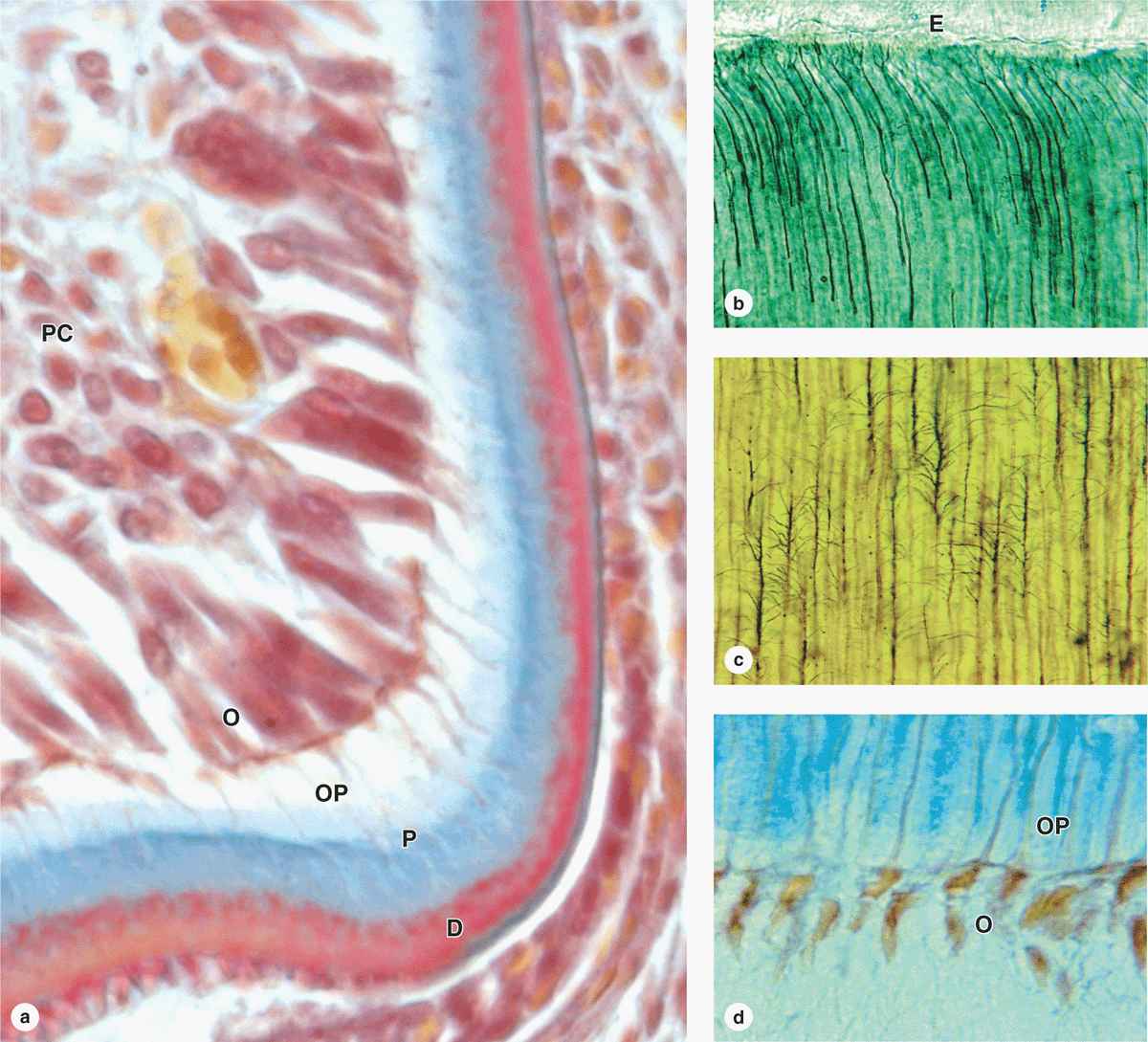
FIGURE 15–7 Dentin and odontoblasts.
Long apical odontoblast processes extend from the odontoblasts within dentinal tubules (Figure 15–7b) that penetrate the full thickness of the dentin, gradually becoming longer as the dentin becomes thicker. Along their length, the processes extend fine branches into smaller lateral branches of the tubules (Figure 15–7c). The odontoblast processes are important for the maintenance of dentin matrix. Odontoblasts continue predentin production into adult life, gradually reducing the size of the pulp cavity, and are stimulated to repair dentin if the tooth is damaged.
Teeth are sensitive to stimuli such as cold, heat, and acidic pH, all of which can be perceived as pain. Pulp is highly innervated, and unmyelinated nerve fibers extend into the dental tubules along with odontoblast processes near the pulp cavity (Figure 15–8). Such stimuli can affect fluid inside the dentinal tubules, stimulating these nerve fibers and producing tooth sensitivity.
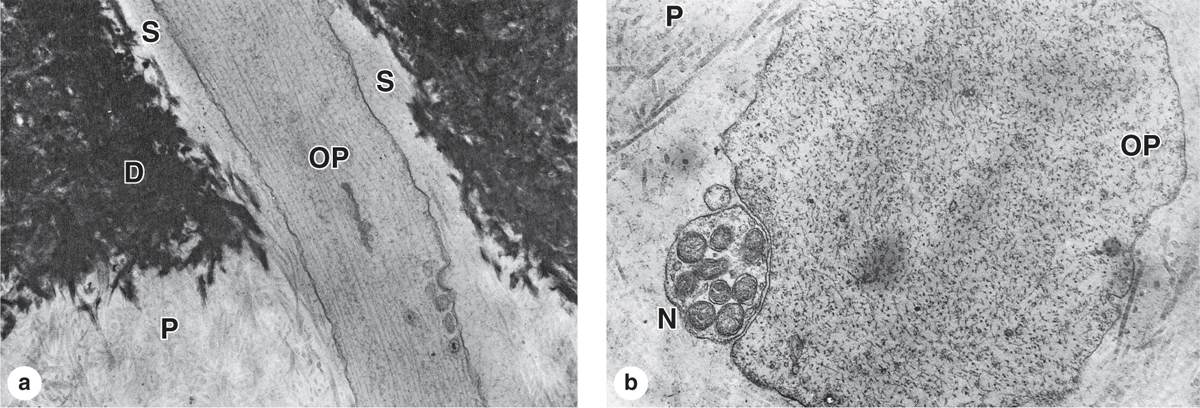
FIGURE 15–8 Ultrastructure of dentinal tubule.
 MEDICAL APPLICATION
MEDICAL APPLICATION
Immune defenses in the oral cavity cannot protect against all infections. Pharyngitis and tonsillitis are often due to the bacterium Streptococcus pyrogenes. White excrescences or leukoplakia on the sides of the tongue can be caused by Epstein-Barr virus. Oral thrush, a white exudate on the tongue’s dorsal surface, is due to a yeast (Candida albicans) infection and usually affects neonates or immunocompromised patients.
Enamel
Enamel is the hardest component of the human body, consisting of 96% calcium hydroxyapatite and only 2% to 3% organic material including very few proteins and no collagen. Other ions, such as fluoride, can be incorporated or adsorbed by the hydroxyapatite crystals; enamel containing fluorapatite is more resistant to acidic dissolution caused by microorganisms, hence the addition of fluoride to toothpaste and water supplies.
Enamel consists of uniform, interlocking columns called enamel rods (or prisms), each about 5 μm in diameter and surrounded by a thinner layer of other enamel. Each rod extends through the entire thickness of the enamel layer, which averages 2 mm. The precise, interlocked arrangement of the enamel rods is crucial for enamel’s hardness and resistance to great pressures during mastication.
In a developing tooth bud, the matrix for the enamel rods is secreted by tall, polarized cells, the ameloblasts (Figure 15-9a), which are part of a specialized epithelium in the tooth bud called the enamel organ. The apical ends of the ameloblasts face those of the odontoblasts producing predentine (Figure 15–10). An apical extension from each ameloblast, the ameloblast (or Tomes) process, contains numerous secretory granules with the proteins of the enamel matrix. The secreted matrix undergoes very rapid mineralization. Growth of the hydroxyapatite crystals to produce each elongating enamel rod is guided by a small (20 kDa) protein amelogenin, the main structural protein of developing enamel.

FIGURE 15–9 Ameloblasts and enamel.
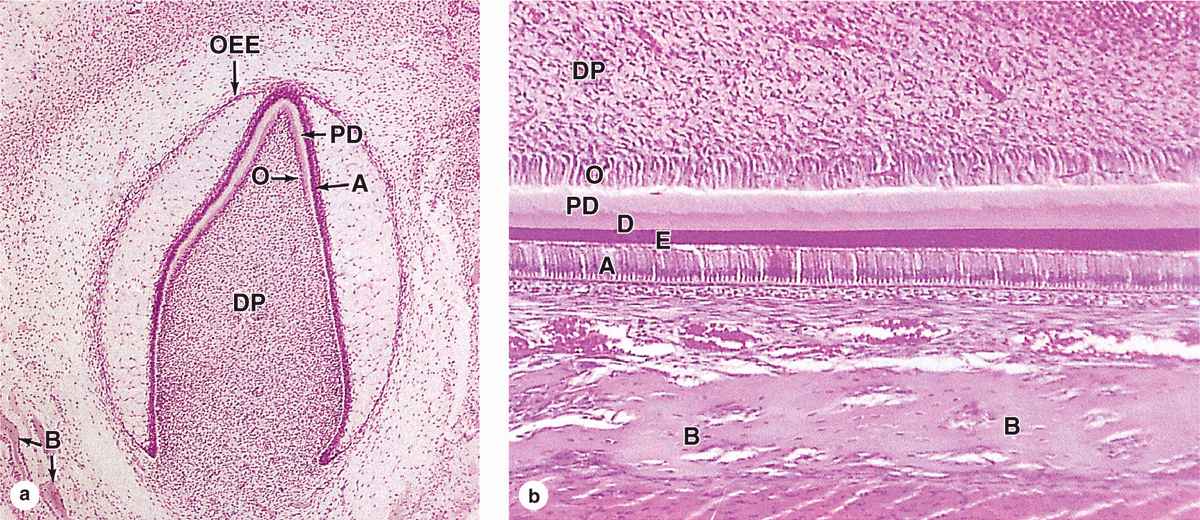
FIGURE 15–10 Tooth formation.
Ameloblasts are derived from the ectodermal lining of the embryonic oral cavity, while odontoblasts and most tissues of the pulp cavity develop from neural crest cells and mesoderm, respectively. Together, these tissues produce a series of 52 tooth buds in the developing oral cavity, 20 for the primary teeth and 32 for the secondary or permanent teeth. Primary teeth complete development and begin to erupt about 6 months after birth. Development of the secondary tooth buds arrests at the “bell stage,” shown in Figure 15–10a, until about 6 years of age, when these teeth begin to erupt as the primary teeth are shed.



