Objectives
- Describe the physiologic and anatomic relationships between the hypothalamus and the anterior and the posterior pituitary.
- Understand the integration of hypothalamic and pituitary function and identify the 2 different pathways used for hypothalamic-pituitary interactions.
- Identify the appropriate hypothalamic releasing and inhibitory factors controlling the secretion of each of the anterior pituitary hormones.
- Differentiate between the routes of transport of hypothalamic neuropeptides to the posterior and anterior pituitary.
- Identify the mechanisms that control the release of oxytocin and arginine vasopressin (AVP).
- Understand the physiologic target organ responses and the cellular mechanisms of oxytocin and AVP action.
The Hypothalamus and Posterior Pituitary Gland: Introduction
The hypothalamus is the region of the brain involved in coordinating the physiologic responses of different organs that together maintain homeostasis. It does this by integrating signals from the environment, from other brain regions, and from visceral afferents and then stimulating the appropriate neuroendocrine responses. In doing so, the hypothalamus influences many aspects of daily function, including food intake, energy expenditure, body weight, fluid intake and balance, blood pressure, thirst, body temperature, and the sleep cycle. Most of these hypothalamic responses are mediated through hypothalamic control of pituitary function (Figure 2–1). This control is achieved by 2 mechanisms: (1) release of hypothalamic neuropeptides synthesized in hypothalamic neurons and transported through the hypothalamo-hypophysial tract to the posterior pituitary, and (2) neuroendocrine control of the anterior pituitary through the release of peptides that mediate anterior pituitary hormone release (hypophysiotropic hormones) (Figure 2–2). Because of this close interaction between the hypothalamus and the pituitary in the control of basic endocrine physiologic function, they are presented as an integrated topic.
Figure 2–1.
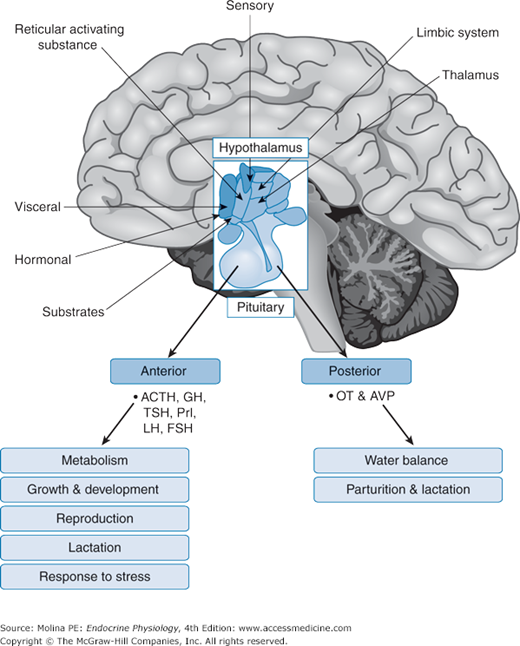
Neuroendocrine regulation of homeostasis. The release of hypothalamic neuropeptides is regulated by afferent signals from other brain regions, from visceral afferents, and by circulating levels of substrates and hormones. The sleep/awake state of the individual, variations in light, noise, fear, anxiety, and visual images all are examples of signals that are integrated by the hypothalamus and that are involved in the regulation of hypothalamic neuropeptide release and control of pituitary function. The hormones released from the anterior and posterior pituitary regulate vital body functions to maintain homeostasis. ACTH, adrenocorticotropic hormone; AVP, arginine vasopressin; FSH, follicle-stimulating hormone; GH, growth hormone; LH, luteinizing hormone; OT, oxytocin; Prl, prolactin; TSH, thyroid-stimulating hormone.
Figure 2–2.
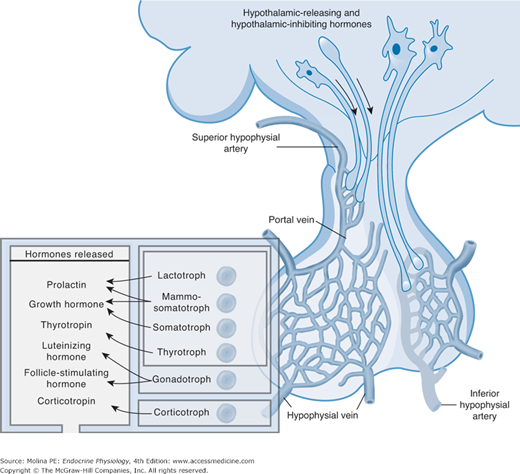
Anatomical and functional relationship between the hypothalamus and the pituitary. The hypothalamus is anatomically and functionally linked with the anterior and posterior pituitary. They are closely related because of the portal system of blood supply. The superior, medial, and inferior hypophyseal arteries provide arterial blood supply to the median eminence and the pituitary. Magnocellular neurons of the supraoptic (SON) and paraventricular (PVN) nuclei have long axons that terminate in the posterior pituitary. The axons of parvicellular neurons terminate in the median eminence where they release their neuropeptides. The long portal veins drain the median eminence, transporting the peptides from the primary capillary plexus to the secondary plexus that provides blood supply to the anterior pituitary. (Adapted with permission from Melmed S. Medical progress: acromegaly. N Engl J Med. 2006;355(24): 2558–2573. Copyright © Massachusetts Medical Society. All rights reserved.)
Functional Anatomy
The hypothalamus is the part of the diencephalon located below the thalamus and between the lamina terminalis and the mamillary bodies forming the walls and the floor of the third ventricle. At the floor of the third ventricle, the 2 halves of the hypothalamus are rejoined to form a bridge-like region known as the median eminence (see Figure 2–2). The median eminence is important because this is where axon terminals of hypothalamic neurons release neuropeptides involved in the control of anterior pituitary function. In addition, the median eminence is traversed by the axons of hypothalamic neurons ending in the posterior pituitary. The median eminence funnels down to form the infundibular portion of the neurohypophysis (also called the pituitary or infundibular stalk). In practical terms, the neurohypophysis or posterior pituitary can be considered an extension of the hypothalamus.
In the hypothalamus, the neuronal bodies are organized in nuclei. These are clusters or groups of neurons that have projections reaching other brain regions as well as ending in other hypothalamic nuclei. This intricate system of neuronal connections allows continuous communication between the hypothalamic neurons and other brain regions. The hypothalamic nuclei can be classified on the basis of their anatomic location or the principal neuropeptide that their cells produce. However, these are not discrete definitions of cell groups; some hypothalamic nuclei may contain more than 1 neuronal cell type. It is best to consider the groups of neurons as clusters of neurons and not as well-defined and delineated nuclei made of a single neuronal type.
Some of the neurons that make up the hypothalamic nuclei are neurohormonal in nature. Neurohormonal refers to the ability of these neurons to synthesize neuropeptides that function as hormones and to release these neuropeptides from axon terminals in response to neuronal depolarization. Two types of neurons are important in mediating the endocrine functions of the hypothalamus: the magnocellular and the parvicellular neurons (Figure 2–3). The magnocellular neurons are predominantly located in the paraventricular and supraoptic nuclei of the hypothalamus and produce large quantities of the neurohormones oxytocin and arginine vasopressin (AVP). The unmyelinated axons of these neurons form the hypothalamo-hypophysial tract, the bridge-like structure that traverses the median eminence and ends in the posterior pituitary. Oxytocin and AVP are released from the posterior pituitary in response to an action potential. Parvicellular neurons have projections that terminate in the median eminence, brainstem, and spinal cord. These neurons release small amounts of releasing or inhibiting neurohormones (hypophysiotropic hormones) that control anterior pituitary function.
Figure 2–3.
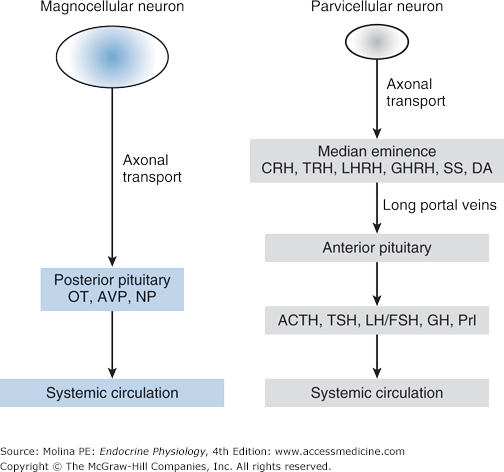
Magnocellular neurons are larger in size and produce large quantities of neurohormones. Located predominantly in the paraventricular and supraoptic nuclei of the hypothalamus, their unmyelinated axons form the hypothalamohypophyseal tract that traverses the median eminence ending in the posterior pituitary. They synthesize the neurohormones oxytocin and vasopressin, which are transported in neurosecretory vesicles down the hypothalamohypophyseal tract and stored in varicosities at the nerve terminals in the posterior pituitary. Parvicellular neurons are small in size and have projections that terminate in the median eminence, brain stem, and spinal cord. They release small amounts of releasing or inhibiting neurohormones (hypophysiotrophic hormones) that control anterior pituitary function will be discussed in the next chapter. These are transported in the long portal veins to the anterior pituitary where they stimulate the release of pituitary hormones into the systemic circulation. ACTH, adrenocorticotropic hormone; AVP, arginine vasopressin; CRH, corticotropin-releasing hormone; DA, dopamine; FSH, follicle-stimulating hormone; GH, growth hormone; GHRH, growth hormone-releasing hormone; LH, luteinizing hormone; LHRH, luteinizing hormone-releasing hormone; NP, neurophysins; OT, oxytocin; Prl, prolactin; SS, somatostatin; TRH, thyrotropin-releasing hormone; TSH, thyroid-stimulating hormone.
The specialized capillary network that supplies blood to the median eminence, infundibular stalk, and pituitary plays an important role in the transport of hypophysiotropic neuropeptides to the anterior pituitary. Hypophysiotropic peptides released near the median eminence are transported down the infundibular stalk to the anterior pituitary, where they exert their physiologic effects. Branches from the internal carotid artery provide the blood supply to the pituitary. The superior hypophysial arteries form the primary capillary plexus that supplies blood to the median eminence. From this capillary network, the blood is drained in parallel veins called long hypophysial portal veins down the infundibular stalk into the secondary plexus (see Figure 2–2). The hypophysiotropic peptides released at the median eminence enter the primary plexus capillaries. From there, they are transported to the anterior pituitary via the long hypophysial portal veins to the secondary plexus. The secondary plexus is a network of fenestrated sinusoid capillaries that provides the blood supply to the anterior pituitary or adenohypophysis. Because of the fenestrated architecture of these capillary vessels, the neuropeptides easily diffuse out of the circulation to reach the cells of the anterior pituitary. The cells of the anterior pituitary express specific cell surface G protein–coupled receptors (see Chapter 1, Figure 1–5) that bind the neuropeptides, activating intracellular second-messenger cascades that produce the release of anterior pituitary hormones.
The blood supply to the posterior pituitary and to the pituitary stalk is provided mostly by the middle and inferior hypophysial arteries and, to a lesser extent, by the superior hypophysial arteries. Short portal vessels provide venous connections that originate in the neural lobe and pass across the intermediate lobe of the pituitary to the anterior lobe. This structure allows neuropeptides released from the posterior pituitary to have access to cells in the anterior pituitary, so that the functions of the 2 main regions of the pituitary cannot be dissociated from each other. Blood from the anterior and posterior pituitary drains into the intercavernous sinus and then into the internal jugular vein, entering the systemic venous circulation.
 As described earlier, 2 general types of neurons constitute the endocrine hypothalamus: the magnocellular neurons, with axons terminating in the posterior pituitary; and the parvicellular neurons, with axons terminating in the median eminence. The neuropeptides released from the parvicellular neuron terminals in the median eminence (corticotropin-releasing hormone, growth hormone–releasing hormone, thyrotropin-releasing hormone, dopamine, luteinizing hormone–releasing hormone, and somatostatin) control anterior pituitary function (Table 2–1). The hypothalamic hypophysiotropic peptides stimulate the release of anterior pituitary hormones. The products released from both the anterior pituitary (adrenocorticotropic hormone [ACTH], prolactin, growth hormone [GH], luteinizing hormone [LH], follicle-stimulating hormone [FSH], and thyroid-stimulating hormone [TSH]) and the posterior pituitary (oxytocin and AVP) are transported in the venous blood draining the pituitary that enters the intercavernous sinus and the internal jugular veins to reach the systemic circulation (see Figure 2–2). Their control and regulation will be discussed repeatedly throughout this book whenever the specific hormone systems to which they belong are described. Several neuropeptides have been isolated from the hypothalamus, and many continue to be discovered. However, only those that have been demonstrated to control anterior pituitary function (hypophysiotropic hormones) and, therefore, play an important role in endocrine physiology will be discussed.
As described earlier, 2 general types of neurons constitute the endocrine hypothalamus: the magnocellular neurons, with axons terminating in the posterior pituitary; and the parvicellular neurons, with axons terminating in the median eminence. The neuropeptides released from the parvicellular neuron terminals in the median eminence (corticotropin-releasing hormone, growth hormone–releasing hormone, thyrotropin-releasing hormone, dopamine, luteinizing hormone–releasing hormone, and somatostatin) control anterior pituitary function (Table 2–1). The hypothalamic hypophysiotropic peptides stimulate the release of anterior pituitary hormones. The products released from both the anterior pituitary (adrenocorticotropic hormone [ACTH], prolactin, growth hormone [GH], luteinizing hormone [LH], follicle-stimulating hormone [FSH], and thyroid-stimulating hormone [TSH]) and the posterior pituitary (oxytocin and AVP) are transported in the venous blood draining the pituitary that enters the intercavernous sinus and the internal jugular veins to reach the systemic circulation (see Figure 2–2). Their control and regulation will be discussed repeatedly throughout this book whenever the specific hormone systems to which they belong are described. Several neuropeptides have been isolated from the hypothalamus, and many continue to be discovered. However, only those that have been demonstrated to control anterior pituitary function (hypophysiotropic hormones) and, therefore, play an important role in endocrine physiology will be discussed.
| Hypophysiotropic hormone | Predominant hypothalamic nuclei | Anterior pituitary hormone controlled | Target cell |
|---|---|---|---|
| Thyrotropin-releasing hormone | Paraventricular nuclei | Thyroid-stimulating hormone and prolactin | Thyrotroph |
| Luteinizing hormone-releasing hormone | Anterior and medial hypothalamus; preoptic septal areas | Luteinizing hormone and follicle-stimulating hormone | Gonadotroph |
| Corticotropin-releasing hormone | Medial parvicellular portion of paraventricular nucleus | Adrenocorticotropic hormone | Corticotroph |
| Growth hormone-releasing hormone | Arcuate nucleus, close to median eminence | Growth hormone | Somatotroph |
| Somatostatin or growth hormone-inhibiting hormone | Anterior paraventricular area | Growth hormone | Somatotroph |
| Dopamine | Arcuate nucleus | Prolactin | Lactotroph |
 Because the hypothalamus receives and integrates afferent signals from multiple brain regions, it does not function in isolation from the rest of the central nervous system (see Figure 2–1). Some of these afferent signals convey sensory information about the individual’s environment such as light, heat, cold, and noise. Among the environmental factors, light plays an important role in generating the circadian rhythm of hormone secretion. This endogenous rhythm is generated through the interaction between the retina, the hypothalamic suprachiasmatic nucleus, and the pineal gland through the release of melatonin. Melatonin is a hormone synthesized and secreted by the pineal gland at night. Its rhythm of secretion is entrained to the light/dark cycle. Melatonin conveys information concerning the daily cycle of light and darkness to body and participates in the organization of circadian rhythms. Other signals perceived by the hypothalamus are visceral afferents that provide information to the central nervous system from peripheral organs such as the intestines, the heart, the liver, and the stomach. One can think of the hypothalamus as a center for integration of the information that the body is continuously processing. The neuronal signals are transmitted by various neurotransmitters released from the afferent fibers, including glutamate, norepinephrine, epinephrine, serotonin, acetylcholine, histamine, γ-aminobutyric acid, and dopamine. In addition, circulating hormones produced by endocrine organs and substrates such as glucose can regulate hypothalamic neuronal function. All of these neurotransmitters, substrates, and hormones can influence hypothalamic hormone release. Therefore, hypothalamic hormone release is under environmental, neural, and hormonal regulation. The ability of the hypothalamus to integrate these signals makes it a center of command for regulating endocrine function and maintaining homeostasis.
Because the hypothalamus receives and integrates afferent signals from multiple brain regions, it does not function in isolation from the rest of the central nervous system (see Figure 2–1). Some of these afferent signals convey sensory information about the individual’s environment such as light, heat, cold, and noise. Among the environmental factors, light plays an important role in generating the circadian rhythm of hormone secretion. This endogenous rhythm is generated through the interaction between the retina, the hypothalamic suprachiasmatic nucleus, and the pineal gland through the release of melatonin. Melatonin is a hormone synthesized and secreted by the pineal gland at night. Its rhythm of secretion is entrained to the light/dark cycle. Melatonin conveys information concerning the daily cycle of light and darkness to body and participates in the organization of circadian rhythms. Other signals perceived by the hypothalamus are visceral afferents that provide information to the central nervous system from peripheral organs such as the intestines, the heart, the liver, and the stomach. One can think of the hypothalamus as a center for integration of the information that the body is continuously processing. The neuronal signals are transmitted by various neurotransmitters released from the afferent fibers, including glutamate, norepinephrine, epinephrine, serotonin, acetylcholine, histamine, γ-aminobutyric acid, and dopamine. In addition, circulating hormones produced by endocrine organs and substrates such as glucose can regulate hypothalamic neuronal function. All of these neurotransmitters, substrates, and hormones can influence hypothalamic hormone release. Therefore, hypothalamic hormone release is under environmental, neural, and hormonal regulation. The ability of the hypothalamus to integrate these signals makes it a center of command for regulating endocrine function and maintaining homeostasis.
Hormones can signal the hypothalamus to either inhibit or stimulate hypophysiotropic hormone release. This control mechanism of negative (or positive) feedback regulation, discussed in detail in Chapter 1, consists of the ability of a hormone to regulate its own cascade of release (see Figure 1–11). For example, as discussed in greater detail in Chapter 6, cortisol produced from the adrenal gland can inhibit the release of CRH, thus inhibiting the production of proopiomelanocortin and ACTH and consequently decreasing adrenal gland synthesis of cortisol. This loop of hormonal control and regulation of its own synthesis is critical in maintaining homeostasis and preventing disease. A shorter loop of negative feedback inhibition also exists, which depends on the inhibition of hypophysiotropic neuropeptide release by the pituitary hormone that it stimulates. In this case, an example would be the ability of ACTH to inhibit CRH release by the hypothalamus. Some neuropeptides also possess an ultrashort feedback loop, in which the hypophysiotropic neuropeptide itself is able to modulate its own release. As an example, oxytocin stimulates its own release, creating a positive feedback regulation of neuropeptide release. These feedback loops are illustrated in Figure 1–12, Chapter 1.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


