The carrier is the archetypal stranger, both embodying the danger of microbial invasion … and transforming it into the possibility for rejuvenation and growth.
–Priscilla Wald in Contagious
Staphylococcus aureus throughout history has been the preeminent scourge of Homo sapiens. Though not a requirement for homeostasis, its ready integration into the flora of the anterior nares and other moist or hairy bodily areas in over 20% of healthy people suggests that S. aureus may function symbiotically at those sites. Yet nasal carriage is the constant element of pathogenesis, the major risk factor for subsequent infection (1, 2and3). After years of tedious delineation of its multiple virulence factors, publications of the whole genomes from heterogeneous strains and new functional studies on regulation and pathogenesis are providing new insights into the mechanisms for invasion of the skin, endovasculature, and solid organs by S. aureus (4, 5and6). Over the last three decades, methicillin-resistant S. aureus (MRSA), because of its relative increase, has overshadowed studies of its relative nonresistant counterpart, methicillin-susceptible S. aureus (MSSA) (7). Yet, MSSA continues to be a major healthcare-associated problem with the risk of MSSA infection in hospital wards three times that of patients who are not carriers (3,8). Much of the phenotypic identity of MRSA comes from the presence of a staphylococcal chromosomal cassette (SCCmec) that with its mecA gene, encodes a new penicillin-binding protein PBP2a, which is necessary for expression of resistance to most beta-lactam antibiotics. Studies of specific genes such as mecA (9) have revealed the complexity of gene expression in pathogenic strains of S. aureus (need new reference). Since the expression of other virulence genes in MSSA and MRSA such as those involving surface adherence is highly regulated like mec, many years of additional study will likely be required to understand pathogenesis in order to design new antimicrobials and vaccines to reduce invasive healthcare-associated infections due to both entities (10, 11and12).
S. aureus as a community pathogen is best known for its ability to produce furuncles and infect soft tissue. A community MRSA known as USA 300 has recently produced a global pandemic, primarily consisting of severe skin and soft tissue infections (13). Along with traditional (USA 200, USA 400) and emerging healthcare-associated strains, USA 300 now threatens hospitalized patients (14,15). The emergence of new healthcare-associated strains comes as no surprise since, historically, healthcare-associated infections were almost exclusively caused by S. aureus until the 1960s, when the prevalence of infections due to gram-negative bacilli increased noticeably (16). That ascent of gram-negative bacteria as the new threat in hospitals lulled hospital physicians into thinking that their old nemesis—S. aureus—would remain of historical interest only. Yet by the early 1990s, data from the National Nosocomial Infections Surveillance (NNIS) system at the Centers for Disease Control and Prevention (CDC) indicated that S. aureus was again increasing in incidence as a healthcare-associated pathogen (17), a trend that only until recently has leveled off (18). With broadening resistance to newer antimicrobials and disinfectants, MRSA along with MSSA has become the dominant healthcare-associated pathogen in hospitals worldwide (19). Additional new strategies are needed to limit healthcare-associated spread and consequent morbidity due to MSSA (20). This chapter reviews the role that MSSA continues to play in healthcare-associated infections and serves as an introduction to Chapter 29, dedicated to MRSA.
HISTORICAL PERSPECTIVE
There are several early biblical descriptions of staphylococcal infection. Of the 10 plagues brought by Yahweh onto the Egyptian Pharaoh, the sixth cast boils or sores upon man and beast (Exodus 9:8-12) (21). The boils arose after Moses took ashes and sprinkled them aloft, filling the air over Egypt with dust that induced outbreaks of boils on man and beast, sores that were wretched in their appearance but not fatal. In another biblical passage, Job is stricken by Satan with boils (or ulcers) that made his body turn black (Job 2:7). There is little information about furunculosis during the next millennium, though it must have remained a major problem. The so-called high Middle Ages has been described as a period that was remarkably disease free, though, ironically, it was followed by centuries of epidemics of plague with little attention to other bacterial infections (22). With the advent of Pasteur’s techniques to culture bacteria, the coagulase-positive Staphylococcus was isolated and assigned a species name in the 1880s (23). Since that time, the number of species of the genus has grown to over 36 (24). Using automated techniques, any clinical laboratory is able to distinguish among the growing number of other species capable of infecting humans (25, 26and27).
Not until the 20th century was a connection made between colonization by specific bacteria and subsequent healthcare-associated infection. The increasing importance of S. aureus as a cause of hospital sepsis resounds from the documentary writing of Dr. Wesley Spink (28). In the preantibiotic era, mortality due to staphylococcal sepsis secondary to pneumonia, osteomyelitis, and cellulitis was as high as 82% (29). Osteomyelitis due to S. aureus, especially infection of the long bones, was often disabling, although mortality due to staphylococcal sepsis was lower in patients with osteomyelitis. The use of sulfonamides from 1937 to 1942, strangely, was not much better than maggots in the treatment of local or osseous staphylococcal infection (28). The miracle of penicillin became available in 1942 and quickly reduced the mortality rate of invasive staphylococcal infection from 80% to 35%. Penicillin resistance, however, developed rapidly in S. aureus, and in hospitals where penicillin was heavily used there were frequent epidemics caused by strains of penicillin-resistant S. aureus (28,30). Multiple advisory groups in the late 1940s, therefore, were assembled to make recommendations for control of staphylococcal epidemics. Pharmaceutical companies, spurred on by the early success of penicillin, mobilized to develop new antistaphylococcal agents (30a).
By the 1950s, Spink et al. established the connection between the carriage in the nasopharynx of hospital employees and the frequent contamination of wounds (31). After penicillin resistance became widespread, Spink’s Minnesota group showed again that the reservoir of S. aureus was hospitalized patients and hospital personnel. Spink stated that the rise in mortality due to penicillin-resistant S. aureus was due to cross-infection of “traumatic and surgical wounds transmitted by healthy hospital carriers or from other patients with sepsis. Patients and hospital personnel were found to be heavily parasitized by highly resistant strains of pathogenic staphylococci primarily within bacteriophage type Group III” (31). Indeed, the problem of staphylococcal sepsis in US hospitals during the early 1950s was the major stimulus for development of infection control committees (16). Such committees, eventually under the direction of hospital epidemiologists, created strict isolation units that, over the next several years, reduced the number of infections at the University of Minnesota and other hospitals. Despite early successes in infection control of staphylococcal infection, Spink’s prescient, cautious words resonate today: “The skin and respiratory tract will remain as the major portals of entry, and staphylococcal sepsis will continue to challenge medical practice.”
Sixty years later, we should remember the intensity of infection control measures that were required for containment of staphylococcal sepsis in the 1950s. Assuredly, during the 1960s and 1970s, newer antibiotics, particularly the semisynthetic penicillins, did reduce the risk of healthcare-associated staphylococcal infection. Extensive use of these agents over the next three decades, however, ushered in international healthcare-associated epidemics due to related MRSA strains (16). In a study comparing the rates of S. aureus infection in a tertiary care hospital from the periods 1971 to 1976 and 1989 to 1992, all but one of the MRSA strains from the later period were acquired in the hospital or in a nursing home, whereas about 80% of the infections due to MSSA were community-acquired (D. Musher, personal communication). Since the third edition of this text was published, the problems of healthcare-associated MRSA and more regularly recognized MSSA with the resultant medical literature have clearly burgeoned worldwide. Over the last 10 years, there have been 8,771 MRSA infections references and 695 MSSA infections references in PubMed, respectively.
MICROBIOLOGY OF STAPHYLOCOCCUS AUREUS
Species Characteristics
Species IdentificationS. aureus has traditionally been defined by phenotypic traits that distinguish it from micrococci and other staphylococci (26). S. aureus is a catalase-negative, coagulase-positive, nonmotile coccus that appears as bluish-black clusters or tetrads after Gram staining. S. aureus grows by 3 days as 6- to 8-mm colonies that are usually hemolytic on blood agar and salt tolerant; they become gold-pigmented after 24 to 48 hours of incubation. Laboratory identification can be aided by observing anaerobic acid production from glucose, production of acid from glycerol in the presence of 0.4 µg/mL erythromycin, and mannitol fermentation, as well as by susceptibility to lysostaphin and furazolidone and resistance to bacitracin (25). S. aureus also produces a thermonuclease that is useful in identifying the species (32). Salt tolerance is probably due to the stability of the S. aureus cell wall derived from the N-acetyl glucosamine residues cross-linked with glycine pentapeptide. Ribitol teichoic acid polymers also link the peptidoglycan (4). Alterations in this structure can actually reduce the minimum inhibitory concentration (MIC) to antistaphylococcal penicillins (33). S. aureus has other survival genes including those to respond to low pH and high salinity, and some of these genes are regulated by sigma factors like σ (34, 35and36).
Staphylococci can be identified by both conventional and rapid laboratory methods. Several marketed kits allow rapid species identification with 70% to over 90% accuracy in detecting as few as 10 CFU/mL (37). Mannitol salt agar has traditionally been the selective culture method of choice for isolation of S. aureus. Newer chromogenic media designed to recover and rapidly identify S. aureus and MRSA appear superior to mannitol salt agar (38). Methods such as restriction fragment length polymorphism (RFLP) of RNA genes (ribotype) and detection of the S. aureus nuclease gene by polymerase chain reaction (PCR) or fluorescein tagging are available commercially to differentiate S. aureus from other species when specialized tracking is required (39). In the future, other methods like MALDI may also become very useful for rapid surveillance of S. aureus healthcare-associated infection/colonization (infra vide).
There are several commercially available PCR assays for use in detecting either S. aureus or MRSA from nasal surveillance swabs. While significantly more expensive than conventional culture methods, several of these PCR assays can be performed on the day of admission to enable colonized patients to be isolated more quickly from the general patient population, thus decreasing the risk of healthcare-associated transmission. In addition, patients undergoing orthopedic or cardiothoracic surgery are often tested for the presence of S. aureus and/or MRSA and decolonized prior to surgery.
S. aureus can be identified directly from positive blood culture bottles by peptide nucleic acid fluorescence in situ hybridization (PNA FISH) or by PCR; the latter method can distinguish MSSA and MRSA. Thus, the healthcare epidemiologist has a growing armamentarium of laboratory methods to expand clinical and epidemiologic investigations.
Strain Identification The healthcare epidemiologist may find it necessary to distinguish among multiple endemic versus epidemic strains of S. aureus for purposes of tracing the source of the infecting isolate, identifying reservoirs of antibiotic-resistant S. aureus, monitoring the colonization of patients or personnel, and, possibly, identifying virulent subtypes. Identification of epidemiologically related microorganisms at the subspecies level may help determine whether the observed clustering of isolates represents distinct strains or several isolates of the same strain that are causing an outbreak. A typing system is adequate if there is a high probability that two random isolates that are epidemiologically unrelated are indeed different. An ideal typing system would generate about 20 groups with even distribution of random isolates. A high proportion of isolates must be typeable, and the method must be reproducible, easy to perform, and inexpensive, and it must avoid the necessity of a second typing system to provide further discrimination. Because of the increasing need to relate strains of S. aureus epidemiologically, methods for typing strains have increased over the last 10 years. Nonmolecular and molecular methods of typing S. aureus are shown in Table 28-1.
TABLE 28-1 Nonmolecular and Molecular Methods of TypingS. aureus
Molecular Methods
Nonmolecular Methods
Plasmid content
Antibiotic susceptibility
Chromosomal REA
Phage typing
Serotyping
Plasmid REA
Capsule typing
RFLP
Alloenzyme patterns
Gene polymorphism
Immunoblotting
Random PCR
Repetitive element PCR
Multilocus enzyme focusing
PFGE
SDS-PAGE
MLST
Whole genome analysis
Microarray analysis
SCCmec type
MLST, multilocus sequence typing; PCR, polymerase chain reaction; PFGE, pulsed-field gel electrophoresis; REA, restriction endonuclease analysis; RFLP, restriction fragment length polymorphism; SDSPAGE, sodium dodecyl sulfate polyacrylamide gel electrophoresis.
Nonmolecular Methods of Typing
Typing systems have been based on differences in antigenic structure, phage susceptibility, antibiotic susceptibility, biochemical profiles, and DNA composition. One antigenic typing method uses antibodies to the 11 known capsular polysaccharides. Healthcare-associated isolates, however, are predominately composed of only two types, 5 and 8 (40), though 24% of strains remain untypeable by this system.
One antigenic system is based on 30 soluble protein or carbohydrate determinants (41). Antigen expression by specific strains depends on the selected growth media, which may be why the antibodies necessary for these systems are not available commercially.
Phage typing has been in use since 1952 and is a laborious method that is performed only by a few reference laboratories. The international set of typing phages, seldom employed but of practical significance, is shown in Table 28-2(16,42). The phage reactions are relatively stable; lysis is graded from weak to strong, and if no lysis develops at 100 times routine test dilution, the strain is considered untypeable. Strains are considered different if the phage pattern differs by two or more phage reactions that show strong lysis. Yet, the same lysis pattern does not necessarily equal epidemiologic relatedness. Phage-typing methods are further limited, because many strains— particularly MRSA—are not lysed by the available phages and are therefore untypeable (42). Phage-typing patterns can change when in vitro selected vancomycin-resistant strains are compared to their vancomycin-susceptible parents (43). Although pulsed-field gel electrophoresis (PFGE) patterns between susceptible parents and resistant selectants remained the same, phage types frequently changed or selectants became nontypeable.
TABLE 28-2 Phages and Phage Groups ofS. aureus
Phage Group
Standard Phages that Lyse
I
29, 52, 52a, 79, 80
II
3a, 3c, 55, 77
III
6, 42e, 47a, 53, 54, 75, 83a, 84, 85
IV
Bovine strains
V
94, 96
Miscellaneous
81, 95
An elegant antigenic typing method, termed immunoblotting, uses pooled human sera for detecting various antigens among S. aureus strains by Western blotting (44). Immunoblotting identified eight patterns using different batches of pooled sera that correlated with phage typing. The method was more discriminating than plasmid profiles, but between 2% and 28% of immunotypes were discrepant on repeat testing. The method was capable of identifying strains that were clinically and epidemiologically related. Immunoblotting combined with antibiograms has been used to discriminate a new outbreak strain from an endemic strain of MRSA (45). Applied in another study, immunoblotting could differentiate 43 strains into only two major groups, making it only as good as endonuclease digestion of plasmid DNA (46).
Antibiograms can be useful when a unique resistance pattern prevails. A given resistance phenotype, however, may result from different arrangements of multiply resistant genes, thus not ensuring DNA sequence identity. Furthermore, resistance (R) plasmids mediating traits such as antibiotic resistance are not always stable. The loss of R plasmids would allow otherwise identical parent strains to be typed as different using antibiograms. A clever way to circumvent this problem is through the use of multiplex PCR that will simultaneously generate multiple amplicons signifying the presence or absence of specific resistance determinants like mecA, aacA, tetM, and so on (47). Multiplex PCR is the current extension of “resistotype” identification suggested by Elek et al. 40 years ago (48).
Biochemical typing (biotyping) is based on enzyme activities, including the commercial methods of such companies as API and Vitek. Color changes are based on acid production from carbohydrates. Although such typing is relatively inexpensive and easy to perform, the traits may vary over time and geographic areas. They are not highly discriminating and need to be combined with other typing methods. Differences in single enzymes, such as esterase, have been used in typing but are also not discriminatory when used alone (49,50).
Multilocus enzyme electrophoresis (MLEE) is based on small differences in electrophoretic mobility of chromosomally encoded metabolic enzymes. Proponents of MLEE feel that each pattern of enzymes determines a clone. MLEE has identified 11 types of S. aureus, though the predominance of type 15 in healthcare-associated outbreaks may limit its usefulness to the healthcare epidemiologist (51). Because it is technically difficult to perform (52), MLEE is also not ideal for hospital outbreaks. Another method analyzes differences in the cell wall peptidoglycan. This method, termed peptidoglycan fingerprinting, separates cell wall components by thin-layer chromatography. Unfortunately, this method is difficult to perform and yields an insufficient number of bands for discriminating among endemic and epidemic strains (53).
In the last 3 years, mass spectrometry has burst onto the clinical microbiology scene as a fast and accurate method for identifying pathogens at a species and even a subspecies level (54). Using one application called matrixassisted laser desorption ionization time-of-flight mass spectrometry (MALDI-TOF MS), species can be obtained quickly in around 90% of instances. Of course, a mass spectrometry instrument is required for local application, so the initial outlay in startup costs is expensive. Nevertheless, recent studies suggest that MALDI-TOF compares to spa typing (infra vide) for rapid and accurate identification of S. aureus clonal complexes (55).
Molecular Typing Techniques
Rapid advances in commercially available DNA manipulation kits, high-throughput sequencing, sophisticated imaging, newer molecular biology instrumentations, etc. have made the molecular genotyping of bacteria feasible in small to advanced clinical microbiology laboratories. DNAbased analysis can differentiate bacterial strains that are similar in phenotypic traits but similar or dissimilar in their genotypes (56). A select number of genotypic methods are described below.
Plasmid Analysis. One of the earliest genotypic methods to distinguish clinical strains of S. aureus was plasmid analysis. Plasmid analysis by either visualization of individual intact plasmid bands in an agarose gel or after digestion with an appropriate restriction enzyme has been a classic genotypic method for more than 20 years. But it has major limitations. First, unlike coagulase-negative staphylococci, S. aureus seldom has more than three plasmids, so there is little chance of generating a complex fingerprint, thus limiting discrimination. Second, as with other bacteria, S. aureus strains may lose plasmids and/or gain or lose antibiotic-resistant genes within a desired plasmid, thus limiting the reproducibility of the method. Third, many strains lack plasmids and are, therefore, untypeable by this method. Fourth, very large plasmids that migrate near the top of the gel may be difficult to differentiate; therefore, precise comparisons between strains with large plasmids would be difficult. In case of large plasmids, inclusion of restriction enzyme digestion steps to digest the plasmid DNA enhances the reproducibility and discriminatory power, though this method is seldom used in clinical microbiological laboratories (57).
Pulsed-Field Gel Electrophoresis. Methods for analyzing chromosomal DNA instead of plasmids overcame many of the limitations of plasmid analysis. PFGE has become one of the most popular methods to genotype S. aureus, particularly MRSA in recent years. In PFGE, chromosomal DNA from S. aureus is digested preferably with SmaI (51,53,58). SmaI digests have been optimized for and shown to be stable in S. aureus strains even after repeated subculture. Since SmaI fragments are too large to permit electrophoresis with a conventional electrophoresis apparatus, a new system has been developed (59). PFGE requires a relatively expensive, specialized apparatus (e.g., CHEF DR II system, BioRad) to resolve the restricted fragments. In PFGE, high molecular weight DNA is extracted in situ from agarose-embedded S. aureus cells using lysostaphin and proteinase K to minimize the shearing of DNA. The in situ DNA (in agarose plugs) is then digested with SmaI, and the digested fragments are resolved in a 1% high-quality agarose gel. During electrophoresis, the direction of the electrical field is alternated to allow the large molecules to be reoriented (60). The alternate or the abrupted electric field enhances the resolution of large DNA molecules (>200 kb and up to 10 MB) by allowing them to snake through the agarose. For S. aureus, the number of SmaI restricted DNA fragments resolved by this method is <20. The similarity and dissimilarity in the fingerprints of two strains are determined by combining genetic relatedness criteria and the Dice Coefficient (61,62): The equation appears as
Sd ={2N × 100}/F,
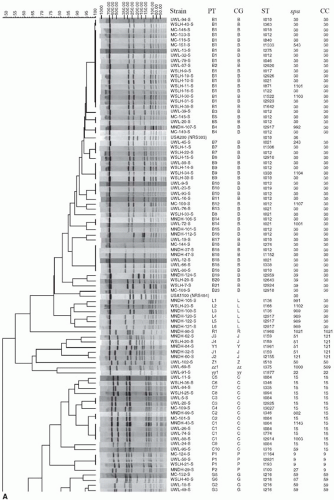
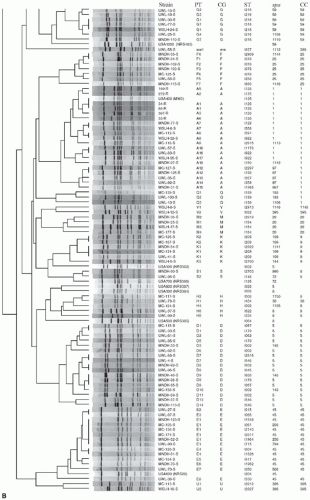
where the Dice coefficient, Sd equals twice N, the number of shared restriction fragments times 100 divided by F, the total number of restriction fragments generated by enzyme digestion (62). Computer software such as Bionumerics is used to establish a DNA similarity matrix based on the Dice coefficient and band tolerance (63,64). The simplicity in interpretation of PFGE results has led it to be the “gold standard” for bacterial genotyping methods. Indeed, PFGE has been extensively used over the last 15 years to describe MRSA outbreaks and their changing epidemiology (58,61,63,65, 66and67). Although PFGE is highly discriminatory and reproducible within a laboratory, methods and technician experience differ among various laboratories and lead to differences that may arise in gel appearance and difficulties when comparing results from different laboratories. Matching of PFGE types through gel images from different sources to implicate spread from one source to another has its limitations. In addition, PFGE is time consuming and takes up to 48 hours to complete the experiment (Fig. 28-1).
Multilocus Sequence Typing. A DNA sequence-based method termed multilocus sequence typing (MLST) has been used for analyzing large collections of bacterial strains from several genera (Table 28-3). This method overcomes the problems with reproducibility and interpretation created by gel-based DNA typing systems. In this method for typing S. aureus, approximately 450-bp long internal fragments of both strands of seven housekeeping genes (carbamate kinase [arcC], shikimate dehydrogenase [aroE], glycerol kinase [glp], guanylate kinase [gmk], phosphate acetyltransferase [pta], triosephosphate isomerase [tpi], and acetyl coenzyme A acetyltransferase [yqiL] are amplified and then sequenced to determine their allelic differences (Table 28-3) (68). The sequence is submitted to the MLST Web site (www.mlst.net) for comparison to known allelic variants to obtain the MLST allelic profile of an isolate. Based on the gene sequence of each of the seven alleles, a sequence type (ST) is determined. Further comparisons between STs can be performed using one of several algorithms. eBURST (electronic Based Upon Related Sequence Types) is one such algorithm. Clonal complexes consisting of related groups or STs are thus generated. This strategy has been used to show marked similarities among a highly diverse collection of MRSA and MSSA (70), reflecting the population and the evolutionary genetics because of the large number of possible STs that can potentially exist. Other advantages of the MLST include its portability between laboratories and availability of a large, freely available, constantly updated MLST database (www.mlst. org/). Unfortunately, the ability to perform PCR and rapid DNA sequencing has limited this method to research laboratories. It should be possible for commercial or university laboratories to adopt the methods and rapidly sequence PCR products generated from epidemiologically important isolates (69). combined the MLST technique with RFLP for tracking genomic islands that may insert into different genetic backgrounds to elegantly show the evolution of MRSA strains from ancestral MSSA strains.
Spa Typing. It was mainly developed to overcome the need of sequencing multiple genes in MLST. Spa typing is based on the polymorphic 21- to 24-bp variable-number tandem repeat (VNTR) within the 3′ coding region of the polymorphic X region of S. aureus-specific staphylococcal protein A. In the spa gene, the in-frame, short-sequence repeats are degenerative, variable in number, and variable in the order in which trinucleotide repeats are organized (70). In this approach, both rapid (microvariation) and slow (macrovariation) genetic variations in the spa gene are indexed. Indexing two types of variations makes it useful in both local and global epidemiologic studies. The sequencing of the polymorphic region allows one to identify the 21- or the 24-bp repeats designated by a letter code (A-Z, A2, B2, etc.), and the profile of these repeats (Y-1-H1-G1-F1-M1-B1-Q1-B1-L1-O1) gives a spa type. A publicly available large database of spa types exists (http://www.egenomics.com/ and Ridom Staphtype software) where one can compare their spa sequence profile to obtain the spa type. The advantages of spa typing are in its lower cost and labor, ease of analysis, and the ever-growing spa database. The disadvantages are that 0.1% of S. aureus strains are non-spa typeable and there is a lack of a strong evolutionary perspective for phylogenetic analysis. Nevertheless, to overcome this shortcoming, BURP (Based Upon Repeat Pattern) has been developed to determine the spa-based clonal complexes (spaCC). The BURP allows the determination of the founder spa types, spaCC, and unrelated types as singletons (71).
Each of the three methods, PFGE, MLST, and spa typing has its own advantages and disadvantages. An investigator needs to know which approach will be most useful for a particular investigation. In general, disease outbreaks investigations are preferably done by the PFGE and now also by spa typing. MLST is the preferred method to study the population structure of the S. aureus and global epidemiology. PFGE has been found to be more discriminatory than MLST and in most cases spa typing as well. One of the main advantages of the PFGE approach is the characterization of major US MRSA clones described as USA100, USA200, USA300 (CA-MRSA), USA400 (CA-MRSA), … USA1200. These USA reference strains are available from the NARSA database (http://www.narsa.net/).
FIGURE 28-1 PFGE dendrogram of S. aureus strains collected over 19 years from Wisconsin and Minnesota. The collection was composed of three groups, including nasal isolates of MSSA, clinical isolates of MSSA, and clinical isolates of community MRSA. Each pulsotype (PT) is also characterized by corresponding PFGE-based clonal group (CG), spa type, and MLST-based sequence type (ST), and clonal complex (CC). Note also the inclusion of PFGE-based USA genotypes that were included as reference strains. The dendrogram was made using 1.25% tolerance, the Dice similarity coefficient, and unweighted-pair group method using arithmetic averages. Strains were considered related if they had >80% genetic similarity. (Reproduced from Shukla SK, Karow ME, Brady JM, et al. Virulence genes and genotypic associations in community-associated methicillin-resistant and susceptible Staphylococcus aureus. J Clin Microbiol 2010;48:3582-3592, with permission.)
FIGURE 28-1 Continued
TABLE 28-3 Housekeeping Genes Amplified by MLST Primers
arc
Carbamate kinase
aro
Shikimate dehydrogenase
glp
Glycerol kinase
gmk
Guanylate kinase
pta
Phosphate acetyltransferase
tpi
Triosephosphate isomerase
yqi
Acetyl coenzyme A acetyltransferase
Multiple-Locus Variable-Number Tandem-Repeat Assay (MLVA). This PCR-based assay exploits the VNTRs in the clfA, clfB, sdrCDE, spa, and sspA loci to obtain PCR amplicon banding pattern that can be used to investigate MRSA outbreaks. (72,73). It is comparable to PFGE if > 75% relatedness for MLVA criteria is used.
Amplified Fragment Length Polymorphism. Amplified fragment length polymorphism (AFLP), which utilizes a combination of selective PCR amplification of restriction fragments from a total digest of genomic DNA, has also been used to study the population structure of S. aureus (74,75). In one study a large collection of S. aureus isolates (n = 1,056), AFLP revealed the presence of three major and two minor phylogenetic branches. The above grouping, however, was not able to distinguish virulent strains from the nonvirulent strains as expected suggesting that any S. aureus genotype could cause a serious infection although some lineages appear to be more virulent than others (75).
Whole Genome Sequencing. Genome sequencing remains the last resort to conclusively genotype a pathogenic strain. Despite the steadily decreasing cost of sequencing, cost is still prohibitive due to personnel time needed in annotation and characterization of novel open reading frames. Yet, with literally hundreds of S. aureus genomesequencing projects in progress, its pan genome features are emerging. The innovative combined approaches of PCR and sequencing will likely spawn a new genechip-based technology that can not only identify different lineages but also distinguish virulent strains from the nonvirulent ones. Host specificity and susceptibility play major roles in the genesis and persistence of staphylococcal infection. This coming decade of investigation should identify the susceptibility markers for S. aureus infection (75a).
Microarray Analysis. A microarray-based approach was published recently that was based on the genome sequence of seven S. aureus genomes and their MLST types. The array was able to identify core genes common to all S. aureus strains besides 10 dominant lineages, although substantial variations were observed in their mobile genetic elements and associated virulence genes (76). These studies did not find any evidence to show that certain S. aureus lineages are associated with invasive isolates in community settings but showed the importance of hypercolonizing strains (77).
Optical Mapping. Optical mapping is a newer tool that relies on the creation of a high-resolution restriction map of a bacterial genome (optical maps) and then comparing with the other optically mapped strains (78). While this is a very powerful technique to visualize major genomic changes with reference to the index or reference genome strains, it is not widely available in clinical microbiology laboratories. It is also limited by the small-sized database of the optical maps.
Optical Fingerprinting by Raman Spectroscopy. Other genotypic tools have been developed in the last few years but are not yet widely available. One novel method has been developed to identify and/or distinguish bacteria clones from the distantly related or the unrelated ones (78a). This method, Raman spectroscopy, works on the principle that Raman Spectra of each bacterial species are a unique clone and therefore could be used to identify the species or cluster them based on their phenotypic properties. Using a small number of S. aureus isolates, it has been shown that this method could be highly discriminatory and may reach the resolving power of PFGE (78a,79).
Summary of Molecular Typing Methods. The three most common methods for genotyping S. aureus are PFGE, MLST, and spa typing. Each of these methods has its own advantages and disadvantages, but there is also a high level of concordance between these three methods. The epidemiologist should ask which genotypic approach will be most useful for a particular investigation. In general, disease outbreak investigations are preferably done by the PFGE because of the ease in interpretation. One of the main advantages of the PFGE approach is the characterization of major US MRSA clones described as USA100, USA200, USA300 (CA-MRSA), USA400 (CA-MRSA), etc. These USA reference strains are available from the NARSA database. Spa typing will likely gain acceptance as sequencing will be more easily accessible in clinical laboratories. MLST is the preferred method to study the population structure and global evolutionary genetics of S. aureus. Shukla et al. (58) showed that the strain-distinguishing ability of spa typing and PFGE were comparable but more discriminatory than MLST for clinical MSSA strains.
PATHOGENESIS OF HEALTHCARE-ASSOCIATED INFECTIONS
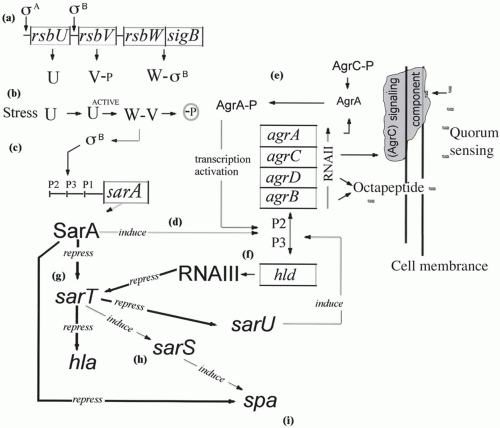
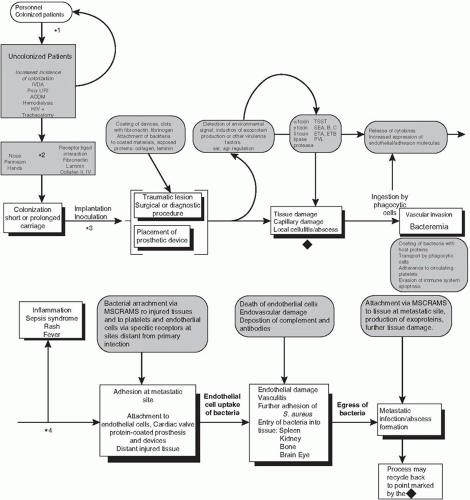
S. aureus is armed with many virulence factors housed on as many as 18 “genome islands.” (80) Most of these virulence factors are highly regulated and are turned on and off depending on the ecological challenge the bacterium faces. Expression of S. aureus attachment and virulence factors results from the interaction of a global accessory gene regulator (agr), a staphylococcal accessory regulator (sar), and RNAIII, a central regulatory function unique to S. aureus (Fig. 28-2). Its regulatory flexibility gives S. aureus opportunity to attach, colonize, and invade many tissues and organs. Because of its invasive properties, S. aureus bacteremia (SAB) has been one of the most studied clinical syndromes (81). In a Brazilian tertiary care hospital, S. aureus caused 21% of all bloodstream infections (BSIs) compared to 26% caused by coagulase-negative staphylocococci (82). Clinical laboratories have observed an increased incidence of bacteremia caused by S. aureus since 1980 (83,84). In a study of healthcare-associated bacteremia of unknown origin, both S. aureus and Pseudomonas aeruginosa caused 15% of the cases (85). S. aureus also caused 50% of catheter-related bacteremias, the majority of infections associated with insertion of prosthetic materials, and the majority of cases of septic arthritis and osteomyelitis. MSSA as well as MRSA strains are extremely prevalent in the intensive care unit (ICU). Among 49 ICUs in Italy, S. aureus caused almost 10% of all ICU infections (86). Understanding the pathogenesis of such a diverse group of infections arising both in the community and in the hospital can be approached by examining the stages of host-pathogen interaction: colonization, attachment, adherence, tissue damage, invasion, dissemination, and metastatic infection (Fig. 28-3) (4).
Colonization
Relationship of Colonization to Infection The major reservoir of S. aureus is the anterior nares. Carriage there influences carriage at other sites, including the axillae, perineum, denuded dermis, and mucous membranes (4,87, 88and89). People who are S. aureus carriers may harbor various strains persistently or intermittently, with intermittent carriage occurring in as much as 90% of a sampled population of carriers (4,90). Some humans, based on genetic and nasal mucus constituents, are noncarriers. Factors that promote colonization include coincident respiratory infection, prolonged hospitalization, needle use (as in intravenous [IV] drug users), diabetics, patients requiring hemodialysis and patients receiving allergy shots, exposure to cold weather, and dermatologic conditions such as eczema (4,91). The elderly, even when they are inpatients, have no higher rate of colonization (92). Antibiotic administration also promotes an ecologic, nasal niche, perhaps through alteration of normal flora that is known to provide resistance to S. aureus colonization (93). Once a patient is colonized with S. aureus, the particular strain may disseminate by personto-person contact, particularly by spread on the hands of personnel (94), or by the dispersion of S. aureus carried on rafts of desquamated skin (95). In this manner, S. aureus strains spread among hospitalized patients. Adhesion factors and global regulators also interact to determine the establishment of colonization (96). Since it usually precedes infection, colonization with S. aureus remains an important risk factor. The importance of the infection of concomitant colonization is shown by the analysis of pathogens in surgical site infections (SSIs) and device-related infections. SSI rates were higher in colonized patients than in noncolonized patients (97, 98and99). This relationship is also quantitative: when the density of colonizing flora exceeds 106 colony-forming units (CFU), rates of postoperative infections are higher among carriers than among noncarriers. Patients in one surgical ICU study who were nasal carriers not only induced cross-colonization but also were significantly more likely to incur a staphylococcal infection than noncarriers (100). Treatment upon admission to the ICU with nasal antistaphylococcal ointment was associated with a lower rate of S. aureus colonization while in the ICU. RFLP patterns were identical for those strains colonizing the nares and causing SSI. Molecular analysis also has been used in clusters of SSIs after heart operations to distinguish between the nasal strain and the strains isolated from SSIs or blood (101).
FIGURE 28-2 Overview of the predicted regulatory pathways involved in gene expression for S. aureus: (a) the sigB operon, transcribed under regulation of τA, encodes rsbU, rsbV, rsbW, and sigB. RsbW is an anti-τB factor that binds to τB, blocking its activity; (b) stress (e.g., high temperature, high osmolarity, or low pH) activates rsbU to U active that then can dephosphorylate rsbV-P to rsbV. RsbV then binds to rsbW, releasing τB; (c) τB binds to a consensus sequence on the sarA P3 promoter, activating transcription of sarA (as well as other promoters); (d) sarA binds to the interpromoter region between P2 and P3 of the accessory gene (agr) locus, stimulating transcription of agr RNAII, which encodes agrB, agrD, agrC, and agrA, elements of a two-component quorum-sensing system. AgrB and agrD produce an octapeptide that diffuses through the membrane to bind to and activate agrC, a membrane-associated signaling component; (e) activated agrC phosphorylates agrA, which induces transcription of agr RNAIII; (f) RNAIII, a pleiotropic regulator for expression of virulence proteins, represses sarT; (g) increased expression of sarT during exponential growth causes repression of sarU, an inducer of RNAIII expression. SarT represses expression of hla (encoding a-hemolysin), and induces expression of sarS; and (h) sarS induces expression of spa (protein A). SarA represses expression of spa (i). (Courtesy of Katherine A. Schmidt and Ambrose Cheung.)
FIGURE 28-3 Pathogenesis of healthcare-associated infection caused by S. aureus. *Potential strategies to interrupt the development of a healthcare-associated S. aureus infection: (1) barrier precautions and decolonization procedures; (2) bacterial receptor (microbial surface components recognizing adhesive matrix molecules [MSCRAMMs]) antagonists and competitive colonization; (3) perioperative prophylaxis, antibiotic impregnated materials, materials to which S. aureus adheres poorly, and active vaccination; (4) prompt institution of effective antimicrobials, cytokine antagonists, MSCRAMM inhibitors, and passive vaccination with monoclonal antibodies; (5) prolonged treatment of bacteremia to cure foci of metastatic infection.
The relationship between nasal carriage and subsequent SSIs is not straightforward. For example, in a study of 414 patients undergoing elective surgery in Khartoum, Sudan, only 6 of the 98 nasal carriers incurred SSIs caused by the strain inhabiting the nose (101a). Besides demonstrating that nasal carriage was not a significant risk factor for development of SSI, the elegant molecular analysis further demonstrated that noncarriers were at significant risk of acquiring an independent SSI caused by strains with a high degree of genetic heterogeneity.
Further implicating S. aureus carriage, the same strains colonizing nonsurgical patients on admission are often the infecting strains (89). Specifically, the same strain colonizing the noses of drug addicts with endocarditis is recovered from blood cultures. Hemodialysis patients are more frequently infected if anterior nares are colonized; 93% of hemodialysis-related infections are caused by the phage type colonizing the anterior nares (102). The same relationship between colonizing and infecting strains was observed for patients on peritoneal dialysis. Nasal carriers had a fourfold higher incidence of dialysis catheter exit site infections (103). It has been estimated that there is a 4% to 16% probability of catheter loss in patients with S. aureus peritonitis who are nasal carriers, compared with a negligible risk for noncarriers (104). Finally, in hospitalized patients, there may be preferential colonization of non-nasal sites, for example, the oropharynx in patients undergoing long-term endotracheal intubation (105). Patients admitted to a large hospital in Meunster, Germany, who developed SAB, were found to be infected 80% of the time with their own nasal-colonizing strain (2). MRSA strains were not a problem in that section of Germany at the time of the study. A larger analysis done in Oxford, England, of the relationship of nasal-colonizing strains to subsequent invasive disease, was conducted by Day et al. (77) using MLST followed by additional analysis using an elegant whole genome microarray analysis for characterizing isolates (76). The major implication of these landmark studies was to show that (106) hypercolonizing community strains mirror the invasive strains and (8) no genes including virulence genes are particularly associated with invasive isolates. Thus, these studies have far-reaching implications for infection control in the community as well as in the hospital.
Most recently, we have further evidence that elimination of nasal strains carried by patients admitted to the hospital will reduce subsequent surgical infections (99,107). In the first study, a randomized, double-blind multicenter trial in The Netherlands, the rate of infection in patients decolonized with nasal mupirocin and chlorhexidine baths was 3.4% compared to 7.7% in patients who remained colonized (relative risk (RR): 0.42; 95% confidence interval [CI]: 0.23-0.75) (99). The second study showed mupirocin decolonization of nasal carriers resulted in a reduction of 6.28 to 3.32 cases per 1,000 patient days (107). These data form the rationale for rapid screening of S. aureus nasal carriers with subsequent decolonization before surgery.
Adherence and Attachment Strains of S. aureus have many potential surface adhesions (Smeltzer book). These adhesions are upregulated by agr once a S. aureus strain contacts a tissue or a surface, a crucial step in the initiation of infection. Adherence is mediated by a group of surface protein adhesins called microbial surface components recognizing adhesive matrix molecules (MSCRAMMs) and regulated by the sar/agr system (108). There are three major groups of MSCRAMMs depending on whether they bind fibronectin, collagen, or fibrinogen, and these are present in strains from asymptomatic carriers (109). Early adhesion is facilitated by upregulation of sar-mediated MSCRAMMs. Furthermore, sarS, a sar homologue, is an activator of S. aureus protein A (110) (Fig. 28-3). Access and adherence to host tissues or implanted materials is mediated by surface receptors that involve host protein interactions. More specifically, the interaction occurs between MSCRAMMS and target structures of the eukaryotic cell. For procedures in which foreign materials are implanted or that result in a thrombus at the surgical site, plasma proteins such as fibronectin are immediately deposited on the materials. Host proteins then act as bridging molecules in the adherence of S. aureus to protein-coated surfaces (111).
S. aureus binding can be blocked by antibodies specific for a particular receptor (6). An additional effect of staphylococcal-protein interaction may allow evasion of the immune system (112). Binding of host proteins to the staphylococcal cell wall effectively coats the bacteria and may prevent host recognition of the microbe. S. aureus may also adhere to uncoated foreign material through electrostatic forces. This interaction is mediated by surface charge and hydrophobicity of the material and bacteria. S. aureus has a net negative charge due to ribitol teichoic acid and protein A (113). Preventionn of infection, however, may require blockade of more than one receptor, a factor that has thwarted development of efficacious staphylococcal adhesion vaccines.
Studies that examine the binding of S. aureus to an extracellular matrix typically use a foreign material such as methylmethacrylate coated with a specific protein. Except for albumin, which markedly diminishes S. aureus binding to polymethylmethacrylate (PMMA), most host proteins augment attachment. For example, S. aureus binding to PMMA is markedly enhanced when fibronectin is present, and this binding is not strain dependent. Fibronectin coats implanted prosthetic material and plastic surfaces and is a major component of the fibrin matrix of clots. Staphylococcal adherence to fibrin clots is increased in the presence of fibronectin (114). Binding to fibronectin is mediated by two related fibronectin-binding proteins, FnBPA and FnBPB, which have specific ligand-binding domains that recognize the N terminal and the C terminal region of fibronectin. When fibronectin coats surfaces, the N terminal end enhances the binding of S. aureus. Fibronectin also binds well to albumin-coated substances and mediates the adherence of S. aureus to collagen, endothelial cells, fibroblasts, platelets, and platelet-fibrin thrombi (115). Strains of S. aureus that bind avidly to fibronectin are more likely to produce endocarditis in a rabbit model of catheter-induced endocarditis, probably because fibronectin is first deposited on valvular endothelial cells traumatized by the catheter (116). In another experiment, fibronectin binding was shown to be crucial in the pathogenesis of endocarditis. Fibronectin-binding deficient mutants were less likely to adhere to damaged heart valves than was the intact parent strain (117).
Similar interactions between S. aureus and other surfaces may also initiate infection. Fibrinogen enhances S. aureus binding to and may be preferentially deposited on IV devices (118,119). Staphylococcal clumping factor has been shown to be the SSP1 that binds to fibrinogen (120,121). Phase 2 clinical trials are underway to assess the therapeutic value of products that block clumping factor (122). Laminin, a major component of the basement membrane, also binds S. aureus by a specific receptor like fibronectin. However, only a small enhancement of S. aureus binding to PMMA was shown (111). Laminin binding may not be a significant factor in the production of intravascular infections, since laminin serum levels are so low. On the other hand, there may be a role for laminin in the production of primary tissue infections, since basement membrane may be exposed after traumatic injury to epithelial surfaces, one of several putative routes that S. aureus can escape the bloodstream (123,124).
S. aureus binds to heparin and other glycosaminoglycans by two bacterial cell wall-associated proteins. Glycosaminoglycans are linked with proteins to form proteoglycans and are found in connective tissue, basement membranes, and eukaryotic cell surfaces. These substances bind to heparinized catheters (125); however, the binding is not specific. Other components of connective tissue bind S. aureus as well. Type IV collagen binds less avidly than fibronectin and laminin to S. aureus (126) but is exposed at the site of tissue injury. Adherence of S. aureus to type IV collagen is enhanced in the presence of fibronectin. In addition, S. aureus binds to type II collagen by a unique receptor, which has been cloned and sequenced. Strains with type II collagen receptors, isolated from patients with osteomyelitis and septic arthritis, were shown to bind well to cartilage (112). Cutaneous injury may promote exteriorization of cytocollagen 10, providing substrate binding not possible in normal skin (126a,127).
Biofilm formation is discussed in Chapter 31 of this book by Fey et al. In brief, the so-called slime substance central to biofilm formation is a polysaccharide composed of beta-1,6-linked N-acetyl glucosamines with partly deacetylated residues. Mutations in the corresponding biosynthesis genes (ica operon) lead to a pleiotropic phenotype wherein staphylococcal cells are less adherent and invasive. Several biofilm-negative mutants have been isolated in which polysaccharide intercellular adhesin production appears to be unaffected (128). Other proteins involved in biofilm formation include accumulation-associated protein (AAP), the clumping factor A (ClfA), the staphylococcal surface protein (SSP1), and the biofilm-associated protein (Bap). New antimicrobials are needed that penetrate and disrupt biofilm formation or that are combined with new polymers to resist adherence and attachment (129). Several biofilm-penetrating antimicrobials as well as biofilm vaccines are under development (130).
Virulence and Invasion
Several regulatory systems that control virulence have been described in S. aureus (Fig. 28-3). The most important of these are sar and agr, both affecting RNAIII, which is a global effector molecule capable of upregulating transcription of many staphylococcal virulence genes. Ironically, staphylococcal binding to platelets also causes release of platelet-bound peptide antibiotics that may ameliorate local infection (4). The production of specific virulence factors by staphylococci results in a complicated cascade of effects depending on the interaction of the regulatory components present. Many of these factors contribute to the antiphagocytic and increased intracellular survival of S. aureus (10). The effects that cause the most severe infections may be produced by strains that harbor a particular complement of regulatory and toxin-encoding genes.
Early events in abscess formation are sar mediated. Subsequent exoprotein and toxin production is regulated by agr. There are at least 34 known exoproteins elaborated by S. aureus and their genetic determinants are grouped in pathogenicity islands throughout the chromosome. Certain proteins are highly toxic and are considered virulence factors. These enterotoxins (A, B, X1,2,3, δ, and E) and toxic shock syndrome toxin-1 (TSST-1) compose a related family of toxins causing staphylococcal food poisoning and toxic shock syndrome and act as superantigens once they enter the systemic circulation (131). Superantigens cause intense activation of certain T-cell populations, and subsequent cytokine production overwhelms the immune system, preventing a coordinated response to antigen processing. The net result of this activation, paradoxically, is similar to endotoxin-induced shock wherein excessive quantities of cytokines induce tissue damage (132,133).
More limited in activity, the epidermolytic toxins (ETAs), exfoliatins A and B specifically, attack the epidermis, causing exfoliation seen in toxic epidermal necrolysis (TEN) and staphylococcal scalded skin syndrome (SSSS). The gene for ETA is located on the chromosome, whereas the exfoliatin B gene (etb) is located on a plasmid. The major pathologic effect of these toxins occurs at a site remote from the site of infection.
Unlike enterotoxins and exfoliative toxins, membranedamaging toxins produce damage at the site of infection. α-Toxin (heat labile), one of four known hemolysins (α, β, γ, δ), is a major pathogenic factor in that it produces tissue damage after the establishment of infection (134,135). Using allelic replacement to create isogeneic toxin-positive and toxin-negative strains, no lesions were generated by the toxin-negative strain in a murine model of mastitis (135). A-Toxin is the only staphylococcal toxin known to damage actively growing nucleated animal cells and is both dermonecrotic and lethal (30 µg/kg) in a murine model (135,136). When α-toxin is injected subcutaneously, vasoconstriction and subsequent tissue ischemia result (30). B-Toxin (a heat-labile sphingomyelinase) and δ-toxin (a heat-stable peptide) are dermonecrotic at high doses but are less potent than a-toxin (134,137). B-Toxin is not produced in many strains because of a converting phage inserted in the hlb gene (136). A staphylokinase (SAK) is carried by the phage and upregulated by agr. Nasal strains usually have SAK intact, whereas SAK-deficient isolates were more than four times as likely to cause a fatal outcome (138).
The δ-toxin peptide is a 26-residue translation product of the hla gene located near the 5′ end of RNAIII encoded by the agr locus. Leukocidin (heat labile), which is toxic to neutrophils and macrophages, also is a potent dermonecrotic toxin (137). It is composed of two proteins, F (32 kd) and S (38 kd), and induces formation of a transmembrane potassium channel (138a). Both components are necessary for toxicity. Leukocidin and γ-toxin belong to the same family of bicomponent toxins (139). The genes for all the dermotoxins (hla, hlb, hld, hlg) are located in the chromosome. One leukocidin, Panton-Valentine leukocidin (PVL), has gained special repute since it is carried by many USA 300 MRSA strains causing community-acquired infection including necrotizing and fatal pneumonia (139a). The extent of the PVL determinant in MSSA strains has not been well studied, but PVL is present in a small number of MSSA isolates (MLST 188) from Malaysia (140).
Other exoproteins, such as proteases, collagenase, hyaluronidase, and lipase, probably act as virulence enhancers and are not as destructive to tissues (136). Although staphylococcal exoproteins are otherwise dissimilar, the expression of at least 12 genes (including α- and β-toxins, exfolitins, enterotoxins B, D, TSST-1, proteases, protein A, and coagulase) is upregulated by agr (141). The agr locus is involved in a two-component regulatory system controlling the expression of virulence genes in other bacteria (142). Other regulatory elements, several of which are sar homologs, have been recently identified (see Fig.28-3). Thus, the complexity of the regulatory elements suggests that virulence factor expression is most likely responding to a variety of environmental and physiologic conditions specific to the host. The agr type and function has recently been related to the mortality associated with staphylococcal bacteremia (143).
Dissemination and Metastatic Infection
After the establishment of local infection, S. aureus may disseminate to other sites. Dissemination from cutaneous sites is infrequent in community-acquired infection but may be more common in healthcare-associated acquisition. Spread of staphylococci from a localized cutaneous infection to the bloodstream and then to deep tissue to form abscesses or to cause endocarditis requires access to the bloodstream and rebinding to potential target sites. In part, S. aureus may gain access to the capillary vascular tree as a result of local inflammation and tissue damage invoked by specific, highly regulated exoproteins. Phagocytic cells may also contribute to vascular entry by carrying viable microorganisms back into the capillaries. Once entry into the bloodstream occurs, binding to serum proteins would follow, and eventually bacteria might stick to a target cell bearing a receptor to either a staphylococcal MSCRAMM or a serum component bound to the staphylococcal cell wall.
S. aureus can also bind to platelets (144), and the binding may increase the capacity of platelets to bind to injured endothelium. Thus, staphylococci may be transported to a distant site and establish a metastatic focus. The ability of bacteria to bind to platelets correlates with the capacity to induce infective endocarditis. S. aureus, with its high capacity to bind platelets, more often caused endocarditis in an animal model than did Escherichia coli, with its minimal platelet-binding capacity. Binding of staphylococci to platelets is direct, rapid, and saturable, suggesting that this property is receptor-mediated and dependent on the number of receptors present. Platelet binding is not dependent on protein A. The staphylococcal ligand is most likely a surface carbohydrate and, perhaps, capsular-based. This ligand is resistant to proteases and susceptible to agents that modify carbohydrates and specific anticapsular antigens (145).
After dissemination, S. aureus must attach to distant tissues to cause a metastatic suppurative infection. Metastatic infections may develop through interactions of blood-borne staphylococci and endothelial cells. These infections may involve endovascular structures or deep tissues. This may be due, in part, to the higher degree of attachment and invasion of endovascular tissue exhibited by staphylococci (146). The interaction of endothelial cells and S. aureus is so efficient that these bacteria adhere to uninjured endothelial cells. Affinity for a specific site usually leads to infection at that site; E. coli microorganisms have been shown to attach to specific uroepithelial cell receptors, and this interaction is a prerequisite for urinary tract infection (UTI) (147). S. aureus appears to have a specific receptor for endothelial cell surface proteins, which promotes adherence and, perhaps, the initiation of endocarditis and graft infections. As with platelet binding, bacteria that bind avidly to endothelial cells are more likely to cause endocarditis than bacteria that bind poorly and consequently rarely cause endocarditis (148).
S. aureus is also a common cause of prosthetic valve endocarditis. Binding of S. aureus to a porcine cardiac valve is a specific receptor-mediated event. In this instance, a binding protein of 120 kd was identified as a potential receptor. This protein was not related to fibronectin. Unlike injured tissues, fibronectin may not augment S. aureus binding to endothelial cells. No fibronectin is expressed on the luminal surface of endothelial cells, and no fibronectin is produced by valvular endothelial cells (149). However, fibronectin may augment the binding of S. aureus to injured endothelial surfaces and if fibronectin binding is blocked, there is a decrease in adherence to subendothelial surfaces exposed after endothelial injury (150,151).
Investigators have also shown that endothelial cells ingest attached staphylococci. A 50-kd protein from umbilical vein endothelial cell membrane binds to S. aureus and facilitates uptake into endothelial cells (152). This protein was shown to be different from fibronectin using a fibronectin antibody assay. Binding is also specific, since albumin or fibrinogen does not inhibit this interaction. Bovine aortic endothelial cells actively phagocytosed S. aureus; 65% of bacteria applied to the endothelial cells were ingested. This action can be blocked by cytochalasin B and was independent of fibronectin and complement (153,154). Complement-activated endothelial cells, conversely, have increased fibronectin binding. Although S. aureus cells that were ingested did not multiply within endothelial cells, the endothelial cells eventually died, leading to exposed subendothelial surfaces. Adherence alone does not induce apoptosis since studies show that viable intracellular S. aureus is needed to induce apoptosis (155). Ingestion and endothelial cell death, however, do depend on strain and inoculum (156). The clinical impact, ultimately, is that intracellular S. aureus may not be affected by most antibiotics, particularly β-lactams, which fail to penetrate eukaryotic cells. Endothelial cell infection would consequently initiate invasion and infection of deeper tissues.
Subsequent interactions between the ingested S. aureus and the endothelial cell may lead to local vascular damage. Uptake of S. aureus by endothelial cells also increases expression of Fc receptors on the cell surface, hypothetically initiating vasculitis by the adherence of neutrophils and platelets to the endothelial cells. Immune complex deposition would next activate complement or initiate the coagulation cascade. These events could augment further metastatic seeding and invasion (157). Finally, host factors elicited by extravascular infection may alter endothelial cells and increase bacterial adherence. For example, staphylococcal adherence to vascular endothelium is upregulated by sar and enhanced in the presence of tissue necrosis factor-a (TNF-a). TNF may further increase the endothelial leukocyte adhesion molecule-1 (ELAM-1) and intracellular adhesion molecule-1 (ICAM-1) (158). Also, the exposure of endothelial cells to lipopolysaccharide increases adherence of bacteria. This effect is duplicated by incubation of cells with the cytokine interleukin-1 (159). Classic agents like aspirin downregulate many pathways associated with complications of endothelial infection and by downregulating global staphylococcal regulons, create novel therapeutic strategies (145).
Specific capsular types of S. aureus play a critical role in abscess induction, as well as in avoiding host phagocytic uptake. Capsules are produced by most clinical strains, and serotypes 5 and 8 together account for up to 50% of clinical isolates (145a).
More than 80% of healthcare-associated isolates from bacteremic patients produce capsule type 5 or type 8 (160). Clinical strains with a type 5 or 8 capsule are more resistant to opsonophagocytosis. In a mouse bacteremia model of infection, a capsule type 5 strain sustained a higher level of bacteremia than two capsule-defective mutants, likely due to the antiphagocytic nature of CP5 since in vitro assays indicated that the parental strain was only susceptible to phagocytic killing by human polymorphonuclear leukocytes (PMNs) in the presence of capsular antibodies and complement. Although capsule types 1 and 2 confer resistance to complement-mediated opsonophagocytosis by PMNs, strains producing these capsule types do not cause clinical disease. CP5 production has also been shown to block adherence of S. aureus to endothelial cells in culture (161). Similarly, in a rat model of catheter-induced S. aureus endocarditis, both the type 5 and 8 parental strains are less pathogenic when compared with capsule-deficient mutant strains (162). These findings suggest that CP5 and CP8 may interfere with staphylococcal attachment to the damaged aortic valve in vivo. Data from mouse models show that a capsule-defective mutant fails to persist in the murine nares (163). Ways to exploit these observations through active or passive vaccination to reduce the risk of healthcare-associated infection await further study.
Only gold members can continue reading. Log In or Register to continue