Small and Large Bowel Structure: Developmental and Mechanical Disorders
STRUCTURE OF THE SMALL AND LARGE INTESTINES
External Appearance
Small intestine. The adult small intestine ranges from 300 to 900 cm in length, depending upon the tone and the degree of stretch induced when measurements are taken. The duodenum constitutes the most proximal portion, is approximately 25 cm in length, and, except for its distal fourth portion, is retroperitoneal and fixed. The duodenum extends from the first portion (duodenal bulb) downward to the second portion on the right side of the spine, to the level of the fourth lumbar vertebrae. The second portion of the duodenum is often referred to as the descending duodenum. The third portion traverses across the spine to complete a C shape that embraces the head and body of the pancreas. The common bile duct and the main pancreatic duct (of Wirsung) usually empty into a common channel, opening at the ampulla of Vater on the medial wall of the descending duodenum. An accessory pancreatic duct (of Santorini) opening is present in up to 70% of patients. It is located 2 to 3 cm proximal to the ampulla of Vater and is the exit for the accessory (minor) duct. The region of the duodenojejunal junction is a fixed landmark fluoroscopically and is referred to as the ligament of Treitz. At this junction, the duodenum becomes unfixed and wrapped in a mesentery.
The proximal 40% of the mobile small intestine is arbitrarily designated as the jejunum and the distal 60% as the ileum. In barium radiographs, the jejunum is concentrated in the left upper quadrant, the jejunoileal region in the midabdomen, and the terminal segments of the ileum in the right lower quadrant. There are some gross differences between the jejunum and ileum. The proximal jejunum is up to 3.5 cm in diameter, or one and one-half to two times as large as the ileum, which measures up to 2.5 cm in diameter. The mesentery of the ileum contains more adipose tissue compared to jejunum. Ileum lies mostly in the lower abdomen and pelvis while jejunum lies in the upper abdomen.
Large intestine. The large intestine comprises that segment of the bowel distal to the ileocecal valve (ICV). It is “large” because of its diameter relative to that of the “small” intestine, the circumference being approximately double. On a daily basis, about 1 L of fluid enters the colon from the ICV, but this is reduced to about 100 mL, which is eliminated. The colon is therefore an absorptive organ for water, electrolytes, bile salt, and other substances produced by bacterial degradation; it also has complex mixing mechanisms as well as acting as a convenient storage facility. It must also resist huge numbers of organisms of different types, some invasive and some toxin producing, as well as a variety of chemicals and potential carcinogens. It begins at the ICV and consists of the right, transverse, and left colon, the transitions between them being at the hepatic and splenic flexures, respectively. The right colon consists of the cecum, which is the saccular part of the large bowel below a horizontal line at the ICV from which the appendix arises, and the ascending colon. The left colon consists of the descending and sigmoid colon together with the rectum.
The large bowel is about 120 to 150 cm in length and characterized externally by its large diameter, the presence of three bands of longitudinal muscle, 0.6 to 1.0 cm in width, running through most of its length, the tenia coli, and the fatty appendices epiploicae. As it passes across the pelvic brim, the descending colon becomes the sigmoid colon, the latter ultimately becoming the rectum at the sacral promontory.
The orifice of the appendix is usually found immediately (1-3 cm) below that of the ICV; the gross and microscopic appearances of the appendix are discussed in greater detail in Chapter 17.
The ascending colon is about 15 to 20 cm in length, running up to the hepatic flexure, at which point it becomes the transverse colon. The latter is 30 to 60 cm in length and is somewhat U shaped, the definition of the U being governed by its length and that of its mesentery, by which it is suspended and dips toward the pelvis. At the splenic flexure, it becomes retroperitoneal and fixed, and passes down toward the pelvis as the descending colon, which is about 20 to 25 cm in length. The sigmoid colon is the natural continuation of the descending colon, beginning approximately where the large bowel crosses the pelvic brim. However, it is also characterized by the presence of a mesentery, although its length, and consequently the length of the sigmoid colon, is very variable, measuring between 20 and 85 cm, with an average of about 40 cm. At the third sacral vertebra, and corresponding roughly to the level of the levator ani muscles at the superior aspect of the internal anal sphincter, the mesentery of the sigmoid colon terminates and the sigmoid colon becomes the rectum. It continues for 10 to 15 cm to the dentate line, which traditionally marks the anatomic anorectal junction. The rectum is longer in men than in women.
The external surface of the large bowel is characterized by the presence of three longitudinally running teniae coli, which are localized condensations of the longitudinal muscle coat and are about 0.5 cm in width. It should be noted that the entire large bowel has a complete external longitudinal muscle coat throughout its length, the teniae being thickenings within this muscle layer. One is close to the attachment of the mesentery, sometimes known as the mesocolic tenia; the other two are antimesenteric, and are almost equidistant from each other and from the mesenteric tenia. One is known as the free tenia (tenia libera), and is easily seen on the anterior surfaces of the ascending and descending colons (Fig. 16-1). The other is the epiploic tenia. It is best seen on the anterior surface of the transverse colon, which folds over anteriorly at the flexures, exposing this tenia. Proximally, the three teniae unite at the base of the appendix, which they completely invest. Distally, at the rectum, all three teniae flare out, forming an external muscle coat similar to that of the small intestine. Blood vessels penetrate the bowel wall on each side of the tenia.
The external surface of the large intestine is also characterized by a series of appendices epiploicae, which are pockets of fat hanging from the bowel, their size correlating with the degree of fat found elsewhere in the body (Fig. 16-1). In the ascending and descending colon, two rows are usually present; in the transverse colon, a single row is found adjacent to the tenia libera on its undersurface. The transverse colon also has the greater omentum hanging from it and the lesser omentum passing up superiorly toward the stomach.
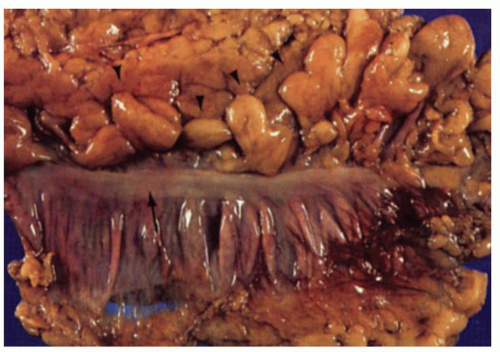 Figure 16-1. External surface of the large bowel showing the free taenia (arrow) and numerous appendices epiploicae (arrowheads). On either side of the tenia, the haustral folds can be seen. |
Peritonealized and Nonperitonealized Surfaces These are of practical importance because carcinomas of the large bowel in these regions can reach both peritonealized and nonperitonealized margins, which assume importance in the staging of carcinomas. Those reaching nonperitonealized margins being pT3, while those reaching (or breaching) peritonealized margins are pT4. The peritonealized margin is not a surgical margin, whereas the nonperitonealized margins often is the surgical resection margin. Inking of nonperitonealized margins in particular therefore should be undertaken to ensure that the relationship between the carcinoma and surgical margin is absolutely clear. At one time, peritonealized margins were called circumferential and the surgical nonperitonealized margins radial, but regrettably these have gradually become used interchangeably, so that now surgical margins are either nonperitonealized, mesenteric, proximal or distal, and nonsurgical margins are peritonealized.
The mesentery is variable along the length of the large bowel, and peritoneum surrounds it variably. Usually, the cecum is completely surrounded by peritoneum and partially mobile. The ascending colon, the hepatic and splenic flexures, and the descending colon are covered only anteriorly and therefore are relatively fixed. Carcinomas of these regions therefore have both peritonealized and nonperitonealized surface. The transverse and sigmoid colon have a mesentery and are therefore surrounded by peritoneum, except at their mesenteric attachments; these latter two regions are therefore particularly mobile. The rectum has no mesentery and rapidly loses its posterior, then its lateral, and finally its anterior covering of peritoneum by its middle third; the lower third is therefore subperitoneal. However, there is marked individual variability in the proportion of rectum covered by mesentery at any particular level. The rectum therefore has no “mesorectum,” simply surrounding tissue, so that the term mesorectal excision is a misnomer, albeit very practical. Of practical importance, and as suggested previously in this section, is that the presence of peritoneum on any surface allows assessment of the adequacy of local tumor excision by direct extension to its margin. In its absence, the irregular resected margins make adequate examination of these margins and comments on the adequacy of local excisions more difficult.1
Once removed from the body, all of these landmarks become almost useless. Any lesion within must be related to the only constant landmarks—the cecal pole, ICV, and pectinate line—or to the resected margins when only part of the large bowel is removed. Externally, the presence of a mesentery immediately suggests the transverse or sigmoid colon, while the flaring of the teniae denotes the rectum. It is therefore always wise for the surgeon to routinely insert a suture into one margin (e.g., proximal), so that the orientation of the specimen is always clear. The greater and lesser omenta are attached to the transverse colon in the immediate vicinity of an adjacent tenia.
Internal Appearance
Small intestine. This is characterized by a series of mucosal folds, which vary from about 3 to 10 mm in height, running transversely around the bowel, although some form spirals running once or twice around the bowel. They are known as valvulae conniventes, although they are neither valves nor close off the lumen, folds of Kerckring or plicae circulares (Fig. 16-2). They gradually diminish through the distal jejunum and may be entirely absent from the distal ileum. These mucosal folds are studded at regular intervals with small nodules, which may vary from pinpoints to 3 mm in diameter and are the sites of underlying lymphoid nodules. The latter become more numerous distally, and in the terminal ileum are represented by larger antimesenteric Peyer’s patches (Fig. 16-2). These lymphoid follicles are very prominent in young individuals and begin
to atrophy in adulthood, so that by old age they may not be visible at all. They are 1 to 1.5 cm in width and 1 to 12 cm in length. They may be more prominent if they become hyperplastic as the result of an underlying disease process, in which historically they are best described, although there are no criteria for lymphoid hyperplasia. Small intestinal villi can usually be identified as pinpoint reflections of light in a freshly resected specimen but can be readily seen using magnification endoscopy. The ICV invaginates the medial wall of the large intestine, forming a large papilla-like structure, the orifice of which folds down to produce a stellate appearance when closed (see Fig. 16-3). The ICV behaves like a physiologic sphincter, preventing coloileal reflux. It consists of two lips, which taper to form folds that are part of the medial cecal wall. In some individuals, there is a very prominent accumulation of fat in the submucosa of the ICV region (Fig. 16-4).
to atrophy in adulthood, so that by old age they may not be visible at all. They are 1 to 1.5 cm in width and 1 to 12 cm in length. They may be more prominent if they become hyperplastic as the result of an underlying disease process, in which historically they are best described, although there are no criteria for lymphoid hyperplasia. Small intestinal villi can usually be identified as pinpoint reflections of light in a freshly resected specimen but can be readily seen using magnification endoscopy. The ICV invaginates the medial wall of the large intestine, forming a large papilla-like structure, the orifice of which folds down to produce a stellate appearance when closed (see Fig. 16-3). The ICV behaves like a physiologic sphincter, preventing coloileal reflux. It consists of two lips, which taper to form folds that are part of the medial cecal wall. In some individuals, there is a very prominent accumulation of fat in the submucosa of the ICV region (Fig. 16-4).
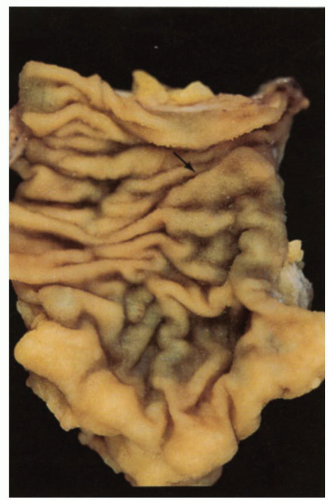 Figure 16-2. Internal surface of the small intestine in which numerous valvulae conniventes (folds of Kerckring) run roughly horizontally across the bowel. At the right center, these are expanded by underlying lymphoid tissue, forming a Peyer’s patch (arrow). Numerous small pinpoints of reflected light on the valvulae represent individual villi that impart a fine granularity to the mucosa. (Courtesy of Dr. J. Frei.) |
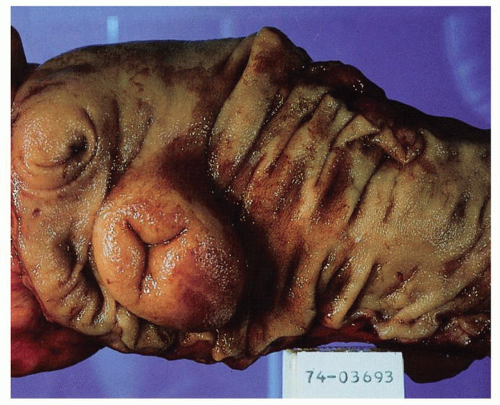 Figure 16-3. Internal surface of the cecum and proximal ascending colon. The ileocecal valve is the most prominent structure, here exaggerated a little by lipohyperplasia. Everything to the left is the cecum, and everything to the right is the ascending colon. The appendiceal orifice can be seen in the cecum. |
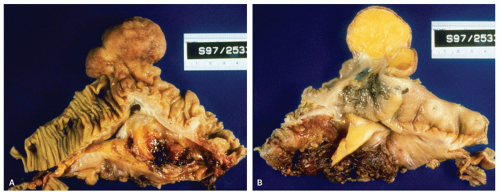 Figure 16-4. Extremely prominent ICV mimicking a tumor (A), that on cut sections shows fat often referred to as “lipoma of the ICV” (B). |
Large intestine. The internal surface of the large bowel is characterized by a series of haustra, which are saccules between the teniae. The boundaries of the haustra are also marked by semilunar folds (plicae semilunaris), which are analogous to the folds of Kerckring in the small intestine but are situated much farther apart and generally cover only part of the circumference (Fig. 16-5). The folds are more prominent proximally and may even disappear or become more irregularly arranged toward the rectum (Fig. 16-5B). The haustra are best seen on imaging studies or during endoscopy and tend to disappear once the colon is resected and opened. Careful inspection of the mucosa also reveals the presence of numerous innominate grooves running primarily transversely across the mucosa every few millimeters (Fig. 16-5D).2 Scanning electron microscopy suggests that between these grooves are smaller furrows into which crypts also open (Fig. 16-5D)3; histologically, these crypts tend to be intimately related focally to underlying lymphoid tissue (see below). These grooves also result in undulations of the mucosal surface forming so-called anthemic folds.
The appendiceal orifice is approximately 2.5 cm (range 1-3 cm) below the ICV, on the medial or posteriomedial cecal wall (Fig. 16-3). It is further described in Chapter 18. Lymphoid nodules can occasionally be identified, particularly in children, in whom they may be slightly dimpled (Fig. 16-5). The wall
thickness of the colon gradually varies from 0.2 to 0.5 cm and increases distally, being thickest in the sigmoid colon.
thickness of the colon gradually varies from 0.2 to 0.5 cm and increases distally, being thickest in the sigmoid colon.
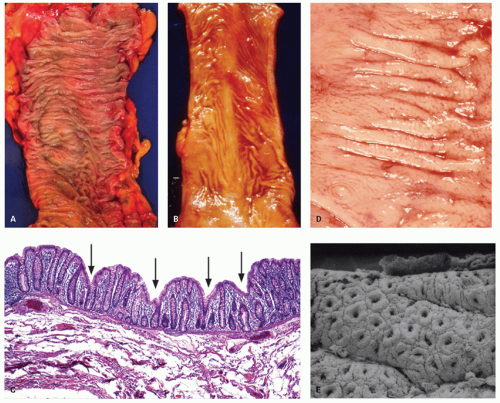 Figure 16-5. A: Mucosal surface of the colon showing multiple transverse mucosal folds (plica semilunaris), and (B) from the rectum showing folds that are somewhat less prominent. The same effect can be produced by stretching the colon after resection. C: Many innominate grooves that are seen as depressions in a well-oriented section of a colonic mucosa (arrows). D: These can also be appreciated on a closer view of the mucosa grossly, which also shows semilunar folds that cover only part of the bowel circumference. E: Scanning electron micrograph of innominate grooves showing three to four crypt openings on their surface between the two grooves shown. |
Although the term rectum is derived from the Latin word meaning straight, because it is the straight part of the intestine (particularly in animals, in which the term was first coined), in practice it has three (and sometimes more) well-defined curves or valves (of Houston), the upper and lower ones curving to the right and the middle one to the left. The upper valve is about 4 cm below the rectosigmoid junction, the lower one about 3 cm above the dentate line. The middle valve is often the largest and is about 6 to 7 cm above the dentate line. It marks roughly the level of the peritoneal reflection and is approximately the limit of the examining finger.
Histology
The layers of the small and large bowels are the mucosa, which includes the epithelium, lamina propria, and muscularis mucosae; the submucosa; the muscularis propria and subserosa or extra rectal fat; and the serosa.
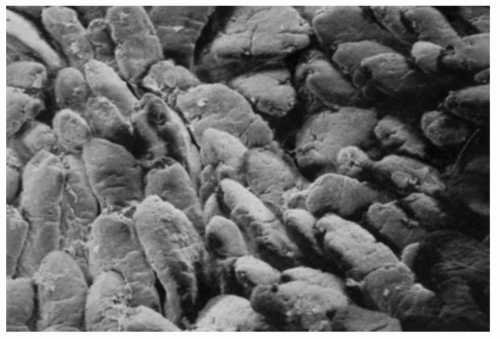 Figure 16-6. Scanning electron micrograph of jejunal villi. Around some of them, horizontal crevices can be seen which are also visible on light microscopy. |
Small intestinal mucosa. The mucosa of the small intestine is lined by finger-like villi 0.5 to 1 mm in height. Scanning electron microscopy shows that the shape of the villi actually varies from being leaf-like to ridges in the duodenum to more finger-like as one moves distally in the jejunum and ileum (Fig. 16-6). The villous height varies in different regions of the small intestine, and in general the villous to crypt ratio varies from 3:1 to 5:1. In the duodenal bulb and descending duodenum, where Brunner’s glands are more abundant, the villi are shorter and stubbier. The villi are tallest in the distal and proximal jejunums and become progressively shorter from the jejunum to the terminal ileum (Fig. 16-7). They have a series of indentations (scalloping) along the sides of the villi, which are marked proximally but may disappear in the terminal ileum (Fig. 16-7). Throughout the small intestinal mucosa, the villi may be distorted by randomly scattered lymphoid follicles. These are much more numerous in the ileal mucosa.
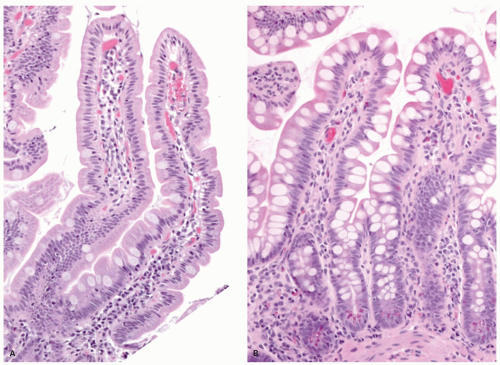 Figure 16-7. Proximal jejunal villi that are tall and leaf-like with scalloping of the borders (A). In comparison the ileal villi (B) appear shorter, more finger-like and contain more goblet cells. |
Epithelium Tall columnar absorptive cells (enterocytes) line the villi. Interspersed among them are goblet cells (Fig. 16-8), which sometimes appear to be increased in the second and third portions of the duodenum, and are unequivocally increased in relation to the number of columnar absorptive cells (enterocytes)
in the ileum. Indeed, a glance at the number of goblet cells is a very easy way of distinguishing proximal from distal small bowel (Fig. 16-7). The nuclei of the villous lining cells are regularly arranged at the bases of the cells. There are scattered intraepithelial lymphocytes in the villi, approximately 1 for every 5 to 10 epithelial cells (Fig. 16-8). Numerous lymphoid nodules are present either as isolated nodules or as aggregates that form Peyer’s patches in the ileum (Fig. 16-9A).
in the ileum. Indeed, a glance at the number of goblet cells is a very easy way of distinguishing proximal from distal small bowel (Fig. 16-7). The nuclei of the villous lining cells are regularly arranged at the bases of the cells. There are scattered intraepithelial lymphocytes in the villi, approximately 1 for every 5 to 10 epithelial cells (Fig. 16-8). Numerous lymphoid nodules are present either as isolated nodules or as aggregates that form Peyer’s patches in the ileum (Fig. 16-9A).
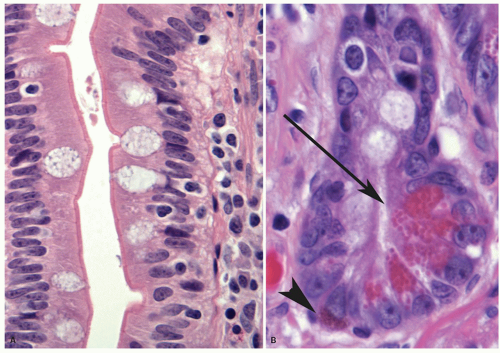 Figure 16-8. A: Close-up of a villus showing the surface epithelium consisting of mostly absorptive enterocytes with few interspersed goblet cells. The surface is lined by a distinct layer of microvilli and glycocalyx that appears as a thick dense line. Scattered intraepithelial lymphocytes are easily seen. B: The base of a crypt in the small intestine that shows many Paneth cells (arrow) characterized by bright eosinophilic granules that are supranuclear, and facing the luminal surface of each cell. An enterochromaffin (argentaffin, Kulchitsky) cell (arrowhead) is also seen near the base that has somewhat smaller subnuclear granules, which are less intensely eosinophilic. Unlike the Paneth cells, the granules are present in the infranuclear portion of the cytoplasm facing the basement membrane, while the nucleus is toward the lumen. |
The predominant cells lining the villi are tall columnar absorptive enterocytes that have numerous microvilli on the luminal surface (Fig. 16-8). On the outer surface of the microvillous membrane is the glycocalyx, a glycoprotein coat just visible on routine hematoxylin and eosin (H&E) sections that can be highlighted by a diastase periodic acid-Schiff (D-PAS) stain or immunostains like carcinoembryonic antigen (CEA) or CD10 antibodies. It is better visualized on electron microscopy as a filamentous coating on the microvillous surface (Fig. 16-10D). This coat is produced by absorptive cells and is firmly adherent to the microvillous membrane. The microvilli of the columnar cells along with villi and mucosal folds (plica circularis) are responsible for providing the extensive surface area needed for absorption.4
The tubules that open at the bases of crypts constitute the crypts (crypts of Lieberkühn). This zone occupies approximately 20% of the total cell column that extends from the tips of the villi to the bases of the crypts. Multiple crypts merge into a vestibule at the base of a villus. In a two-dimensional plane, that is, in welloriented sections, one or two crypts are seen in continuity with some villi. The most abundant cells in the crypts are relatively undifferentiated cells that contain small cytoplasmic granules not usually visible at the light microscopic level. There is no distinct landmark that separates crypts from villi. In light microscopic sections, a band is seen approximately one-fifth of the way up from the muscularis mucosae, and this is often taken arbitrarily as the approximate crypt-villous junction. The columnar cells and goblet cells from the villous
surface are in continuity with the crypt epithelium. The crypts in addition harbor the undifferentiated stem cells and Paneth cells. Mitoses are seen near the crypt bases, with an average of one to two mitotic figures per welloriented crypt section. Paneth cells are present at the base of the crypts, and each crypt contains about 5 to 12 Paneth cells (Fig. 16-8B). The crypts also contains goblet cells, as well as cells that appear intermediate between undifferentiated crypt cells and goblet cells. The goblet cells that line the villi and crypts resemble those found elsewhere in the gut. Endocrine cells are also scattered throughout the crypt zone and are less frequently distributed along the sides of villi (Fig. 16-8B).
surface are in continuity with the crypt epithelium. The crypts in addition harbor the undifferentiated stem cells and Paneth cells. Mitoses are seen near the crypt bases, with an average of one to two mitotic figures per welloriented crypt section. Paneth cells are present at the base of the crypts, and each crypt contains about 5 to 12 Paneth cells (Fig. 16-8B). The crypts also contains goblet cells, as well as cells that appear intermediate between undifferentiated crypt cells and goblet cells. The goblet cells that line the villi and crypts resemble those found elsewhere in the gut. Endocrine cells are also scattered throughout the crypt zone and are less frequently distributed along the sides of villi (Fig. 16-8B).
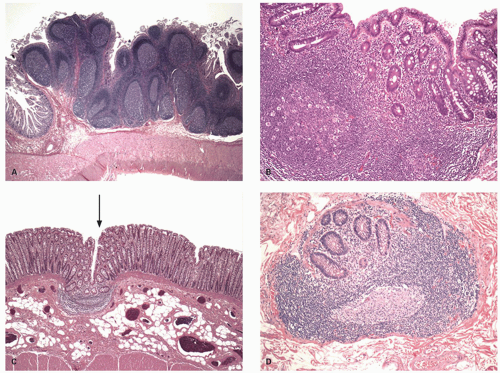 Figure 16-9. A: Section of the terminal ileum showing normal Peyer’s patch. These can become very large in children or under immune stimulation, and can grossly mimic tumors. Note that even at this magnification the villi on top of the lymphoid follicles appear shorter compared to those away from it. B: The lymphoglandular complex in the colon showing the reduced number of goblet cells in the vicinity of the lymphoid aggregate. The crypts seem to be oriented obliquely, which may create an impression of crypt distortion in biopsies. C: The innominate groove (arrow) showing presence of a lymphoid follicle that extends into the submucosa. D: The lymphoid complexes along with crypts may extend in the submucosa, sometimes when cut tangentially may result in confusion with invasive glands in biopsies. |
Lamina Propria The lamina propria is the connective tissue core of the villi and extends from the villous tips to the muscularis mucosae. The most numerous cell types within the lamina propria are mononuclear cells, primarily plasma cells, and lymphocytes. Plasma cells are concentrated more in the intercrypt region. Other cell types in the lamina propria are much more sparsely distributed including eosinophils, macrophages, and mast cells. Normally neutrophils are rare. Longitudinal and branched eosinophilic fibers in the lamina propria of the villi represent smooth muscle cells that can be mistaken for macrophages (Fig. 16-11). Noncellular connective tissue elements, that is, collagen and elastin, are also present. Small blood vessels and fibroblasts and lacteals are in close contact with the basal laminae of the absorptive cells. The central lacteal is usually not easily seen unless pathologically enlarged or highlighted by immunohistochemical stains; this is probably the result of starvation before biopsy or resection. In the
duodenum and distal ileum, villi are normally stubbier and thus appear to contain increased lamina propria mononuclear (plasma cell and lymphocyte) cells compared to the mucosa from other regions of the small bowel.5 Numerous fine nerve twigs are present in the lamina propria, many of which pass through, or invest mast cells.
duodenum and distal ileum, villi are normally stubbier and thus appear to contain increased lamina propria mononuclear (plasma cell and lymphocyte) cells compared to the mucosa from other regions of the small bowel.5 Numerous fine nerve twigs are present in the lamina propria, many of which pass through, or invest mast cells.
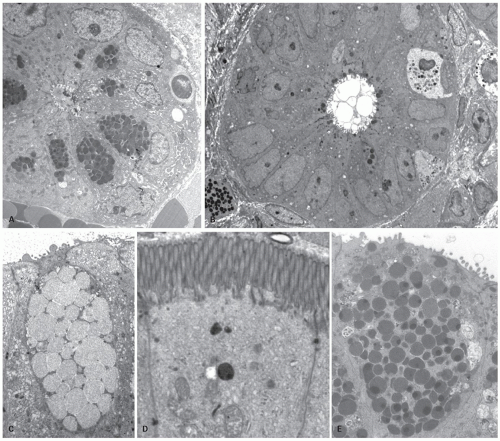 Figure 16-10. Electronmicroscopy showing cross section of a base of a crypt in small and large bowel. A: Small bowel crypt showing many Paneth cells and few cells lack granules and likely represent undifferentiated (stem) cells. B: Cross section of a crypt base from colon showing many undifferentiated cells and some with few Paneth cell type granules. Higher magnification to show the ultrastructure of a goblet cell containing many mucin vacuoles (C), absorptive enterocyte with microvilli and glycocalyx (D). E: Paneth cells with numerous large supranuclear granules. |
ILEAL PIGMENT. Very frequently, a fine black pigment can be seen in macrophages in ileal biopsies, which is most prominent in the deep lamina propria or superficial submucosa, (Fig. 16-12) or within the macrophages in the region of Peyer’s patches. The pigment has been shown to contain calcium, phosphorus, titanium dioxide and, aluminum, and magnesium silicates. It is thought to be derived from diet, possibly from substances such as toothpaste which may contain titanium dioxide as a whitening agent; however, the mechanism speculative.6, 7 It appears to have no clinical significance, although when very prominent can cause diagnostic issues, especially if the patient has a history of melanoma. A similar pigment can be seen in the duodenum, often very prominently in some cases when it is referred to as melanosis duodeni or more appropriately psuedomelanosis duodeni, as the pigment is not melanin (Fig. 16-13). The pigment has
shown to be ferrous sulfide. Pseudomelanosis duodeni has been associated with chronic renal failure, use of antihypertensive drugs, iron pills, and prior mucosal hemorrhages.7, 8
shown to be ferrous sulfide. Pseudomelanosis duodeni has been associated with chronic renal failure, use of antihypertensive drugs, iron pills, and prior mucosal hemorrhages.7, 8
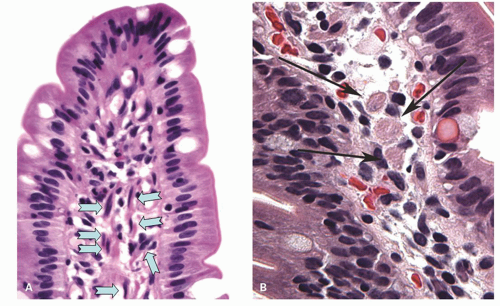 Figure 16-11. A: Villus showing longitudinal smooth muscle fibers (blue arrows), with many lymphocytes and plasma cells in the lamina propria. A few intraepithelial lymphocytes are easily recognized. B: Another villus sectioned tangentially showing transversely sectioned reddish smooth muscle fibers that can be confused with lamina propria macrophages. |
Ileocecal valve. The gross appearance of the aperture of the ICV is described above, and histologically also, many variations can occur.9 Usually a band of smooth muscle (Fig. 16-14A-C) that is contiguous with the muscularis propria of colon and ileum is seen running through the center of the valve and seems to end up joining the muscularis mucosae right at or very close to the junction of ileal and colonic mucosa (Fig. 16-14E).10 In children, its lips may be thickened because of the quantity of lymphoid tissue present, and the valve may appear everted in some individuals whereby the nodularity is visible from the colonic side (Fig. 16-14C). In adults, occasionally these nodules are biopsied as suspected colonic polyps. The adipose tissue in the substance of the valve varies among individuals and can be very prominent in some. This lipohyperplasia may mimic or be indistinguishable
from a true lipoma and is often even referred to as lipoma of the ICV (Fig. 16-4).
from a true lipoma and is often even referred to as lipoma of the ICV (Fig. 16-4).
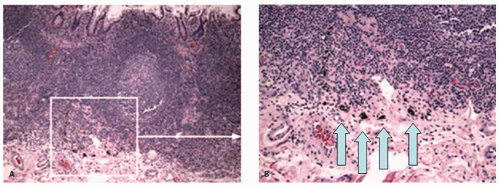 Figure 16-12. A: Ileal biopsy with lymphoid aggregate and foci of a dark brown to black pigment predominantly in the macrophages, but also present extracellularly. Inset (B) shows close-up of the pigment in the macrophages. This is not an uncommon finding in ileal biopsies in the areas of Payer’s patches and has no clinical significance. |
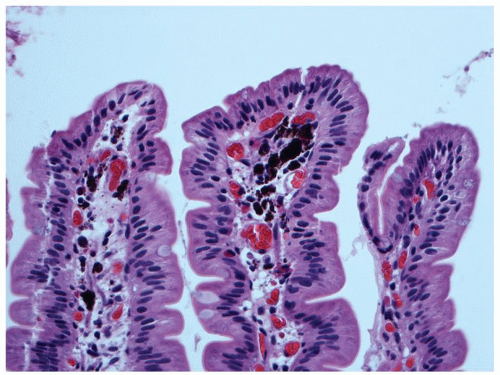 Figure 16-13. Pseudomelanosis duodeni showing accumulation of dark black pigment in the lamina propria macrophages in the duodenal mucosa. |
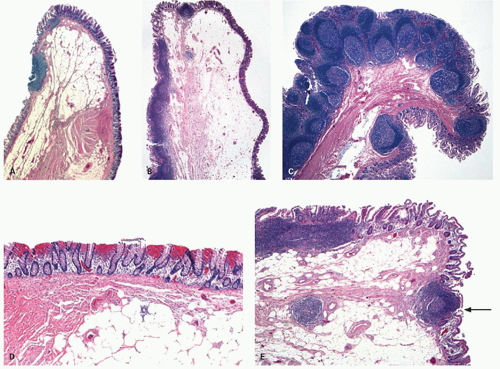 Figure 16-14. Section of ICVs showing normal variations. A-C: Each ICV shows a thick band of muscle in the center that attaches to the muscularis mucosa at the junction of ileal and colonic mucosa. Notice adipose tissue normally present in the ICV. The ileum shows a prominent Peyer’s patch that may protrude through the ICV (C) and be biopsied during colonoscopy as a polyp. D: The colonic mucosa in this junctional area often shows mild inflammatory changes and crypts that are less than perfectly oriented with regards to the surface, something that should be kept in mind while interpreting biopsies from ICV. E: The central muscular band in the ICV that attaches to muscularis mucosa end separates the ileal and colonic sides. |
Large intestinal mucosa. Although the luminal surface of the colon is considered to be smooth, other than for the presence of haustra, careful examination of the surface mucosa reveals a series of surface irregularities that tend to go transversely around the bowel and are reminiscent of the small lines of fingerprints. These innominate grooves are readily seen using a hand lens or, even better, by low-power scanning electron microscopy (Fig. 16-5).3, 11 Similar but less well-defined lines join parallel irregularities to form small geographic areas. When this configuration is examined using ordinary light microscopy, the same changes are apparent, although there is some variation from patient to patient and even from area to area within the same patient. Stretching the mucosa
when pinning out resected specimens prior to fixation tends to make these changes less apparent and inevitably raises the question of whether they are normally present or represent an artifact. The fact that they are readily seen even with the naked eye if looked for, and are also visible both radiologically2 and even in biopsy material, strongly suggests that they are part of the normal anatomy. When viewed using the scanning electron microscope, it is apparent that the crypt openings are far better defined on the left colon than on the right.3 The reason for this is not clear.
when pinning out resected specimens prior to fixation tends to make these changes less apparent and inevitably raises the question of whether they are normally present or represent an artifact. The fact that they are readily seen even with the naked eye if looked for, and are also visible both radiologically2 and even in biopsy material, strongly suggests that they are part of the normal anatomy. When viewed using the scanning electron microscope, it is apparent that the crypt openings are far better defined on the left colon than on the right.3 The reason for this is not clear.
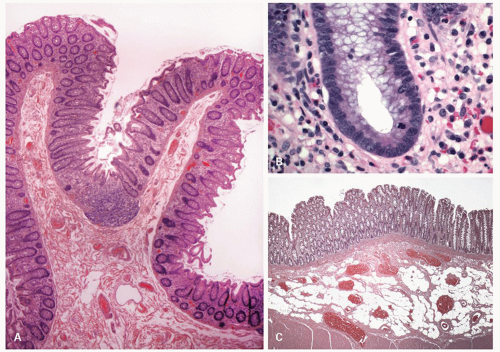 Figure 16-15. A: Overview showing colonic mucosal folds (plica semicircularis). Also note that at the bottom of the fold the crypts dip into a lymphoid aggregate similar to that seen in Fig. 16-9C. B: Detail of the base of colonic crypt showing lack of goblet cells, a population of undifferentiated cells in which the stem cells reside, and a mitotic figure. This is the zone that is occupied by the progenitor cells. C: Submucosal adipose tissue in colonic resection that is an incidental finding of no clinical significance. |
On light microscopy, it is apparent that crypts open both onto the surface of these ridges (anthemic folds) and into the crevices (innominate grooves) formed by them (Figs. 16-15 and 16-16). The function of the crypts in these grooves is likely as an antigen sampling apparatus analogous to the M-cells in the small intestine, and to increase the surface area, which may facilitate absorption, as does the presence of villi in the small intestine. However, careful examination by light microscopy, particularly if sections are not embedded perfectly, shows that at the base of many of these crypts is a lymphoid aggregate (Fig. 16-9C). Further examination reveals a change in the nature of the epithelium in the immediate vicinity of these aggregates from one that primarily contains goblet cells to one that appears to
consist entirely of absorptive cells (Fig. 16-9B). It is also apparent that there is an intimate relationship between the aggregate and the overlying follicle, with close apposition of one to the other but also with numerous lymphocytes passing through the epithelium, the majority of which are T cells (see subsequent discussion on lamina propria and immune system).12 Thus they are often referred to as lymphoglandular complexes rather than merely lymphoid aggregates.
consist entirely of absorptive cells (Fig. 16-9B). It is also apparent that there is an intimate relationship between the aggregate and the overlying follicle, with close apposition of one to the other but also with numerous lymphocytes passing through the epithelium, the majority of which are T cells (see subsequent discussion on lamina propria and immune system).12 Thus they are often referred to as lymphoglandular complexes rather than merely lymphoid aggregates.
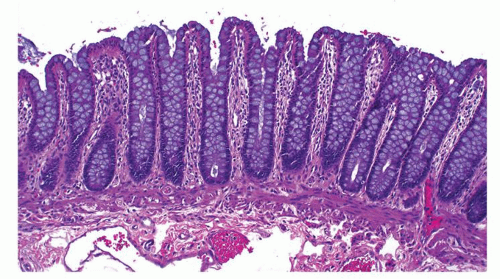 Figure 16-16. Normal colonic mucosa showing perpendicularly oriented crypts parallel to each other like test tubes in a rack. Note that the crypt bases almost touch the muscularis mucosae and are full of goblet cells, except at the crypt bases. The crypts at the edges of anthemic folds, which correspond to the innominate grooves, seem to converge at the base creating a false impression of crypt branching as seen here. The muscularis mucosae have two layers of smooth muscle consisting of an inner circular and outer longitudinal layers that can be appreciated here, but may not be always obvious. The lamina propria shows a scant amount of normal lymphoplasmacytic infiltrate. |
It is little less than amazing that a single layer of epithelium separates the potentially harmful contents of “bad outside world” in the bowel lumen from the internal milieu of the “body beautiful.” However, this protection is provided by a combination of protective mucus rich in secreted immunoglobulin A (IgA) that entraps bacteria, peristalsis that moves this layer of mucus, various antimicrobial secretions from the Paneth cells, and other component of the innate and adaptive immune system. This epithelium, besides forming a protective layer, performs a variety of tasks, reflected in the specialization into different cell types. The two predominant cells lining the colonic crypts are absorptive colonocytes (columnar) and goblet cells. The former are indistinct in the crypts where they are barely visible other than for their nuclei, while goblet cells are most conspicuous. The absorptive enterocytes on the other hand predominate on the surface where absorption occurs (Figs. 16-15 and 16-17) within the crypts. The absorptive colonocytes are tall columnar cells that show microvilli and a glycocalyx layer on the luminal surface similar to intestinal absorptive cells. They also contain special mucin vesicles that are different from goblet cell mucin. The apical portion of these cells fans out covering the tops of the adjacent goblet cells, such that only small part of the goblet cell opens into the lumen (Fig. 16-10). A common artifact seen in colonic biopsies at a low magnification is the appearance mimicking thickened subepithelial collagen layer (Fig. 16-17). On closer look this artifact is due to lighter staining of the infra-nuclear portion of the cytoplasm and the nucleus being pushed up in the middle of the cell. A fair number of endocrine cells are present through out the colon, and specialized microfold (M) cells are largely present in the areas of the lymphoid complexes. Paneth cells may normally be found on the right side, particularly in the vicinity of the ICV and the proximal ascending colon (Fig. 16-18). Paneth cells are generally absent beyond hepatic flexure, although rarely can be seen in the proximal transverse colon.
Specialized cells in the small and larger bowel epithelium
Paneth Cells Paneth cells have a very distinctive appearance with large bright eosinophilic granules clustered near the luminal aspect of the cells (Fig. 16-9B). Some cells that are more sparsely granulated and are intermediate between Paneth cells and goblet cells have also been recognized. Paneth cells have a relatively long life span of 20 days compared to 3 to 5 days for enterocytes. These cells have now been shown to play an important role in body’s defense against microbial infection and play an important role in innate and adaptive immune responses.13 They have been shown to contain a variety of antibacterial peptides and proteins such as lysozyme, alpha-defensins, secretory phopholipase-2, cryptdin-related sequence peptides, and angiogenin-4. Paneth cells secrete these antibacterial products at high levels in response to cholinergic stimuli and when exposed to bacterial antigens.13, 14 The release of these products into the crypt lumen is suggested to protect adjacent mitotically active crypt cells that are responsible for renewal of epithelium against enteric infections. The Paneth cells express a variety of toll-like receptors (TLRs) and nucleotide-binding oligomerization domain-2 (NOD2), both of which play an important role in innate immune responses in GI tract.13 TLRs are transmembrane molecules, while NOD proteins are cytosolic and recognize microbial components after their invasion into the cell. These belong to a group of pattern recognition receptors that are germ line coded and do not require prior exposure to the pathogen. They recognize highly conserved motifs designated “pathogen-associated molecular patterns” (PAMPs), which are present in many microorganisms, but not in their hosts. These motifs are considered essential for the pathogenicity and survival of the microbes. Activation of these pattern recognition receptors (TLRs and NOD2) leads to release of various anti-microbial agents and chemokines that recruit other cellular component of the immune response. Thus, Paneth cells likely serve as guardian or protectors of the mucosa generally, but also the precious stems cells population located at the crypt bases. Mice transgenic for a human Paneth cell alpha-defensin, HD-5 are completely immune to infection and systemic disease from orally administered Salmonella enterica serovar typhimurium.15 In another experimental model, it has been shown that newborn mice are susceptible to infection by Shigella, but become resistant to infection by day 7 when the Paneth cells develop.16 Experimentally depleting the mice of Paneth cell induces the susceptibility to the same infection.16 These evidences provide a strong evidence for the role of Paneth cells in resistance to enteric infections. In humans, lack of Paneth cells in newborn infants has been linked to development of neonatal necrotizing enterocolitis (NEC).17 It is believed that lack of lysozyme may render these infants susceptible to bacterial translocation and subsequent sepsis. More interestingly, the Paneth cells have been now linked to inflammatory bowel disease (IBD), particularly Crohn’s disease.13 NOD2 recognizes a constituent
of peptidoglycan of bacterial cell wall called muramyl dipeptidase. Mutations in the NOD2 gene located on chromosome 16 have been associated with a group of patients with Crohn’s disease, whereby the defective gene product fails to recognize bacterial muramyl dipeptidase, leading to increased susceptibility of the intestinal mucosa to invasion by luminal bacteria and development of chronic inflammation. On the other hand, Paneth cell metaplasia in the colon in various chronic inflammatory conditions including IBD may be an attempt to protect the damaged epithelium from luminal microbes. Decrease in Paneth cells has also been reported with development of complications in celiac sprue (refractory celiac sprue and ulcerative jejunoileitis/enterpathy-associated T-cell lymphoma).18
of peptidoglycan of bacterial cell wall called muramyl dipeptidase. Mutations in the NOD2 gene located on chromosome 16 have been associated with a group of patients with Crohn’s disease, whereby the defective gene product fails to recognize bacterial muramyl dipeptidase, leading to increased susceptibility of the intestinal mucosa to invasion by luminal bacteria and development of chronic inflammation. On the other hand, Paneth cell metaplasia in the colon in various chronic inflammatory conditions including IBD may be an attempt to protect the damaged epithelium from luminal microbes. Decrease in Paneth cells has also been reported with development of complications in celiac sprue (refractory celiac sprue and ulcerative jejunoileitis/enterpathy-associated T-cell lymphoma).18
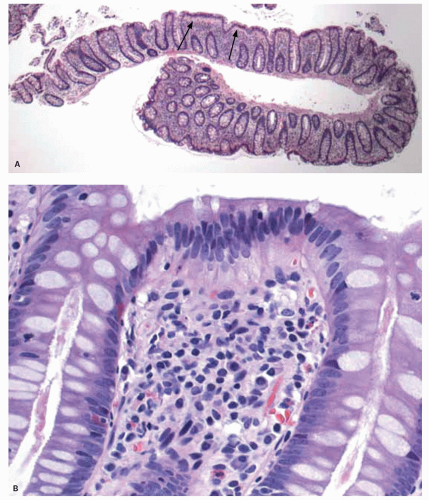 Figure 16-17. A: Overview of a colonic biopsy showing a common artifact that mimics thickened subepithelial collagen (arrows). B: Detail shows that the artifact is due to paler staining of the infra-nuclear portion of the cytoplasm of the surface enterocytes. |
Endocrine Cells Endocrine cells are also scattered throughout the crypt zone in both small and large bowel, and are less frequently distributed along the sides of villi. They can be demonstrated with a variety
of techniques including silver stains, IHC markers and electron microscopy. Morphologically, two types are recognized. One type has inconspicuous granules on H&E sections, often communicates with the lumen, and has a basal nucleus. These cells cannot be easily identified on H&E sections. The other type is the enterochromaffin cell with its characteristic subnuclear orange granules that are either pyramidal or spindle shaped on H&E sections (Fig. 16-8). In the small intestine, more gut endocrine cells are located in the upper small intestine than in the lower portion, and rare endocrine cells can sometimes be found apparently isolated in the lamina propria. The endocrine cells throughout the intestinal tract can be easily highlighted using either silver stains or immunostains. The silver stains identify two types of cells. The most easily demonstrable of these are argentaffin cells that stain the serotonin-producing enterochromaffin cells. The name argentaffin implies that they will cause the deposition of silver from ammoniacal silver nitrate onto themselves in the absence of another reducing agent; the Masson-Fontana stain has precisely these characteristics. By contrast, other endocrine cells require a reducing agent for silver to be deposited—a so-called argyrophil stain; examples include the Grimelius, and Chirukian and Schenk stains (Fig. 16-19). At least three other types of argyrophil cells containing densecore neurosecretory granules can be demonstrated.19
Immunohistochemical stains identify at least 30 different types of endocrine cells distributed throughout the GI tract. Immunostains, because of their higher specificity and sensitivity, are superior to the silver stains, and have replaced them in practice for the identification of endocrine cells. The various cells identified in the intestines include cells that contain serotonin, somatostatin, cholecystokinin, secretin, gastric inhibitory peptide, enteroglucagon, substance P, neurotensin, vasoactive intestinal polypeptide (VIP), enteroglucagon, substance P, L cells, and motilin. Regional differences in the distribution of these cells exist. It is interesting that adenomas and adenocarcinomas of the large bowel not infrequently contain Paneth cells and endocrine cells. However, so-called goblet cell carcinoids (usually appendiceal) contain goblet cells, neurosecretory cells, and rarely Paneth cells, while true carcinoid tumors are almost entirely endocrine. This likely suggests origin of the adenomas and carcinomas from a more primitive cell type compared to endocrine tumors. It is interesting that in the midgut (jejunum, ileum, and proximal large bowel), virtually all endocrine tumors are of the enterochromaffin cells type. Distal to this in the large bowel, L-cell tumors occur, and the distribution of the tumors mirrors the distribution of these cells in the large bowel, which predominate in the rectum, although these tumors are also common in the appendix. This admixture of neurosecretory cells with all other cell lines in neoplasms, together with the elegant work of Le Douarin,20 strongly indicates that these endocrine cells in the GI tract are of epithelial (endodermal) and not neural crest origin.
of techniques including silver stains, IHC markers and electron microscopy. Morphologically, two types are recognized. One type has inconspicuous granules on H&E sections, often communicates with the lumen, and has a basal nucleus. These cells cannot be easily identified on H&E sections. The other type is the enterochromaffin cell with its characteristic subnuclear orange granules that are either pyramidal or spindle shaped on H&E sections (Fig. 16-8). In the small intestine, more gut endocrine cells are located in the upper small intestine than in the lower portion, and rare endocrine cells can sometimes be found apparently isolated in the lamina propria. The endocrine cells throughout the intestinal tract can be easily highlighted using either silver stains or immunostains. The silver stains identify two types of cells. The most easily demonstrable of these are argentaffin cells that stain the serotonin-producing enterochromaffin cells. The name argentaffin implies that they will cause the deposition of silver from ammoniacal silver nitrate onto themselves in the absence of another reducing agent; the Masson-Fontana stain has precisely these characteristics. By contrast, other endocrine cells require a reducing agent for silver to be deposited—a so-called argyrophil stain; examples include the Grimelius, and Chirukian and Schenk stains (Fig. 16-19). At least three other types of argyrophil cells containing densecore neurosecretory granules can be demonstrated.19
Immunohistochemical stains identify at least 30 different types of endocrine cells distributed throughout the GI tract. Immunostains, because of their higher specificity and sensitivity, are superior to the silver stains, and have replaced them in practice for the identification of endocrine cells. The various cells identified in the intestines include cells that contain serotonin, somatostatin, cholecystokinin, secretin, gastric inhibitory peptide, enteroglucagon, substance P, neurotensin, vasoactive intestinal polypeptide (VIP), enteroglucagon, substance P, L cells, and motilin. Regional differences in the distribution of these cells exist. It is interesting that adenomas and adenocarcinomas of the large bowel not infrequently contain Paneth cells and endocrine cells. However, so-called goblet cell carcinoids (usually appendiceal) contain goblet cells, neurosecretory cells, and rarely Paneth cells, while true carcinoid tumors are almost entirely endocrine. This likely suggests origin of the adenomas and carcinomas from a more primitive cell type compared to endocrine tumors. It is interesting that in the midgut (jejunum, ileum, and proximal large bowel), virtually all endocrine tumors are of the enterochromaffin cells type. Distal to this in the large bowel, L-cell tumors occur, and the distribution of the tumors mirrors the distribution of these cells in the large bowel, which predominate in the rectum, although these tumors are also common in the appendix. This admixture of neurosecretory cells with all other cell lines in neoplasms, together with the elegant work of Le Douarin,20 strongly indicates that these endocrine cells in the GI tract are of epithelial (endodermal) and not neural crest origin.
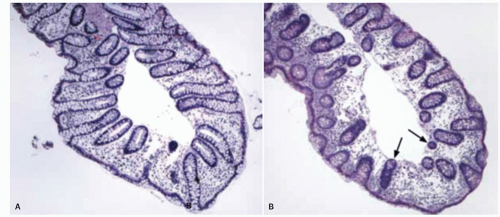 Figure 16-18. A: Biopsy from the left side of the colon showing normal histology. B: Biopsy from the right side of the colon from the same patient, which is also normal but shows denser lymphoplasmacytic cells in the lamina propria and occasional Paneth cells. A branching crypt likely represents two crypts opening at the base of an innominate groove. |
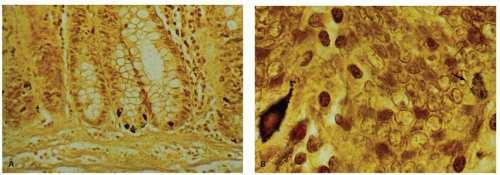 Figure 16-19. A: Grimelius stain showing several dark-staining endocrine cells primarily in the crypt base. B: Detail shows a strongly staining cell (right), which in a parallel section is an argentaffin (enterochromaffin) cell. On the left is a much more faintly staining argyrophil cell (arrow), the precise identity of which depends primarily on immunoreactivity with specific antibodies and occasionally on electron microscopy. |
Goblet Cells The cells are named so because of their shape that resembles wine goblets. The mucous globules in the goblet cell cause the luminal aspect of the cytoplasm to balloon, and the nucleus appears cramped and scalloped at the base of the cell. In animals, variants of columnar absorptive cells, but with shortened microvilli that appear preferentially to attract bacteria, have been described as cup cells while other cells have microvilli extending beyond those of normal enterocytes, so-called tuft cells.21 They have yet to be adequately documented in man. The goblet cells strongly stain with Muc2, a mucin that has a protective affect against luminal infectious organisms.22, 23, 24 It has been calculated that goblet cells make up 20% to 30% of the cells in the right colon and 40% to 50% of those in the left colon. There may be a gradual increase in the number of goblet cells from the right colon to the left, but this pattern is rarely clear enough to allow one to predict reliably from which part of the large bowel a particular biopsy specimen was taken. Nevertheless, scanning electron microscopy and mucin staining (Fig. 16-20) also suggest that there are differences between both sides of the large bowel. There is also a change in the quality of mucus on both sides of the colon, particularly the ratio of sulfated to nonsulfated mucin,25 and lack of sulfated mucin has been correlated with possible premalignant changes.26, 27, 28 Alterations in mucin structures in cancer have many biologic and pathologic consequences.29 They can influence the growth and survival of the caner cells, their ability to invade and metastasize, and their interactions with lectins and cell-surface receptors or cells of the immune system. Mucins also interact with adhesion molecules of cells and can block cell-to-cell adhesion mediated by integrins and E-cadherin. Mucins can also inhibit cell lysis by natural killer cells and interactions with cytotoxic lymphocytes.
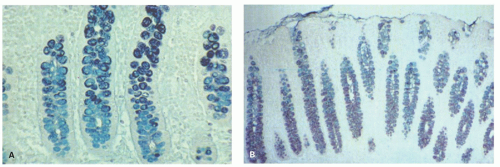 Figure 16-20. High-iron diamine alcian blue stain from the right and left colons. A: In the right colon, dark-staining sulfomucins predominate in the upper part of the mucosa, while in the left colon (B) the converse is the case. |
The mucins covering the surface not only provide a physical barrier, but allow adherence to many commensal bacteria that competitively inhibit binding of the pathogenetic organisms. The secretion of mucins can be modified by activation of pathogen recognition receptors (PRR) and TLR present on the epithelial cells.22
Microfold (M) Cells M cells are so called because of their surface microfolds (microfold cells) or thin membranous cytoplasm (membrane cell) rather than microvilli. The cells are membrane-like (and also macrophage-like), rather than the adjacent tall columnar epithelium. They are not readily visualized by light microscopy and lack a specific immunohistochemical marker for easy identification, although in rabbits they express vimentin that helps in differentiation from adjacent enterocytes.30 They have been largely characterized by electron microscopy.4, 31
These cells typically have poorly organized brush border with fewer microvilli and lack a glycocalyx layer.32 Many lymphocytes can be seen indenting the cell cytoplasm, almost appearing as if they are within the cell cytoplasm. In many places, a thin sheet of cytoplasm is all that separated the lymphocytes from the lumen; M cells transport soluble and particulate antigens from the lumen to the cells in the underlying lymphoid follicle and can therefore be regarded as the component of gut-associated lymphoid tissue (GALT) epithelial cells, or more generically called mucosaassociated lymphoid tissue (MALT); however, they also transport macromolecules in the opposite direction.33 They transport the luminal antigens, various macromolecules, bacteria, or virus to the lamina propria to be exposed to the immune system. Whether these cells can also present antigens to the immune system similar to dendritic cells is still unclear. Large particles and bacteria are internalized via phagocytosis, while viruses and adherent particles by endocytosis via clathrin-coated vesicles.32 Surprisingly, they are also characterized by the absence of secretory component (SC) expression in the animal model34; they therefore appear to be unable to secrete IgA, which may, at least partially, explain the sensitivity of these cells to a variety of organisms and their toxins. While most of the M cells are present in the vicinity of the lymphoid follicles, they can be seen in the surface epithelium of villi as well as elsewhere away from the lymphoid tissue.35 It is still unclear whether the M cells are derived from transformation of adjacent enterocytes, although recently it has been suggested that they are possibly derived from a separate pathway from the progenitor cells.30
Stem cells and kinetics. The mature cells near the tip of the villi or surface of the colon die from apoptosis and are continuously either shed into the lumen, or, especially in the right side of the large bowel, undergo apoptosis. As the cells die at the surface, transient gaps or holes are created that are immediately sealed off by adjacent cells. Apoptotic nuclei are often ingested by macrophages immediately beneath the surface epithelium that may be lightly pigmented. Cell replication within small and large bowel occurs in the crypts and with a turnover time of about 3 to 6 days. Due to this high turnover of cells in the small and large bowel, it is not surprising that the progenitor cells constitute an important cell population in the crypts. Their role in cancer development has further ignited a significant interest in them.36, 37 Most of the data come from experiments in mice, and ingenious labeling studies have helped to track the pattern of cell proliferation and movement of cells in the epithelium, especially in the small bowel.38, 39 It has been fairly clear for many years that the stem cells are located at the base of the crypts although the precise location and identification has been debated. The most actively dividing cells, as seen with long-term tritiated thymidine incorporation, bromodeoxyuridine (BRDU) incorporation or proliferation markers (Ki67) (Fig. 16-21), are seen at the sides of the crypts just above the level of Paneth cells and have a feedback loop with the adjacent Paneth cells (Fig. 16-22). These are called transit amplifying cells and undergo rapid proliferation producing about 300 cells per crypt per day that eventually differentiate into four major intestinal cell types-absorptive enterocytes, goblet cells, endocrine cells, and Paneth cells. However, it appears that the stem cells are fewer and reside at the deepest part of the crypt base and are surrounded by Paneth cells on each side.37 These have been called crypt base columnar cells (CBC).40 It is now shown that these stem cells express Lgr5/Gpr49 gene, which encodes an orphan G protein-coupled receptor and appears to be reliable stem cell marker in the intestine. The stem cells are believed to undergo asymmetric division by which they produce one stem cell (self-renewal) and another committed cell that become committed progenitor cells and undergo further divisions to differentiate into various other intestinal cells types. Under normal conditions, Ki67 stain shows only occasional positive nuclei at cyrpt bases in the stem cells zone (Fig. 16-21C); however, under stress or conditions that require rapid cell renewal, the zone expands.
The cells arising from these progenitor cells migrate upward in the crypts and eventually on to the luminal surface or surface of the villi, a process that takes approximately 3 to 6 days in man. As the cells move upward, they differentiate into absorptive cells, goblet cells, M cells, and endocrine cells. The cells migrating downward into the crypts differentiate into Paneth cells, which move to the bottom of the crypts. The differentiation of the cells into various cell types is complex and involves NOTCH and Wnt signaling pathways. The progeny from a single stem cell at the bottom of a crypt represents a clone of cells that moves up along the crypt in the form of a ribbon. Sometimes the entire crypt may consist of a single clone.
Pericrypt fibroblast sheath. Surrounding the crypts of the small and large bowel is the pericrypt fibroblast
sheath, which is closely apposed to the epithelial cells (Fig. 16-23). In some biopsies, this is relatively prominent, in others virtually invisible. A complex epithelial-mesenchymal interaction exists between these pericryptal fibroblasts and the crypt epithelium. The fibroblasts have been thought to proliferate at the same rate as the crypt epithelium and to migrate with the epithelium.41 However, some studies suggest that the fibroblast may in fact have a much slower turnover time, which would necessitate the epithelial cells “sliding” up along the sheath.42 This fibroblastic sheath is absent around the gastric glands, and it has been shown in a study that with intestinal metaplasia in the stomach with the development of intestinal type epithelium, a pericryptal fibroblastic sheath also develops.43 While it may therefore be better to think of the crypts as an integral epithelial-fibroblast complex, this concept poses problems. If both crypts and fibroblasts migrate at the same rate, what happens to the fibroblasts when they reach the epithelium immediately beneath the lumen? Clearly, they do not simply pile up and form tumor masses; possibly they escape into the lumen by passing through or between surface epithelial cells. Presumably, they either die immediately and are phagocytosed by local macrophages, transform into another cell population (possibly subepithelial macrophages), migrate elsewhere, or are simply not formed in the same numbers as epithelial cells.
sheath, which is closely apposed to the epithelial cells (Fig. 16-23). In some biopsies, this is relatively prominent, in others virtually invisible. A complex epithelial-mesenchymal interaction exists between these pericryptal fibroblasts and the crypt epithelium. The fibroblasts have been thought to proliferate at the same rate as the crypt epithelium and to migrate with the epithelium.41 However, some studies suggest that the fibroblast may in fact have a much slower turnover time, which would necessitate the epithelial cells “sliding” up along the sheath.42 This fibroblastic sheath is absent around the gastric glands, and it has been shown in a study that with intestinal metaplasia in the stomach with the development of intestinal type epithelium, a pericryptal fibroblastic sheath also develops.43 While it may therefore be better to think of the crypts as an integral epithelial-fibroblast complex, this concept poses problems. If both crypts and fibroblasts migrate at the same rate, what happens to the fibroblasts when they reach the epithelium immediately beneath the lumen? Clearly, they do not simply pile up and form tumor masses; possibly they escape into the lumen by passing through or between surface epithelial cells. Presumably, they either die immediately and are phagocytosed by local macrophages, transform into another cell population (possibly subepithelial macrophages), migrate elsewhere, or are simply not formed in the same numbers as epithelial cells.
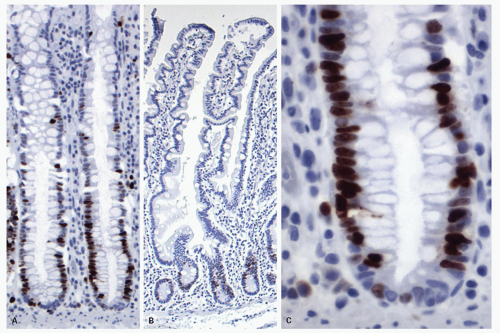 Figure 16-21. Ki-67 staining to show aggregation of positive nuclei at the crypt bases showing the location of the proliferative compartment (progenitor cells) in the colon (A)
Get Clinical Tree app for offline access
Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree


|