21 FIG. 21.1 Taste buds FIG. 21.2 Olfactory receptors FIG. 21.3 The eye (illustration opposite) FIG. 21.4 Eye, monkey FIG. 21.5 Wall of the eye FIG. 21.6 Retina FIG. 21.7 Retinal photoreceptors FIG. 21.8 Neuronal interconnections in the retina FIG. 21.9 Fovea
Special sense organs
Introduction
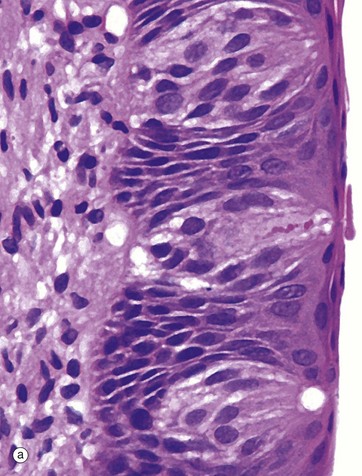
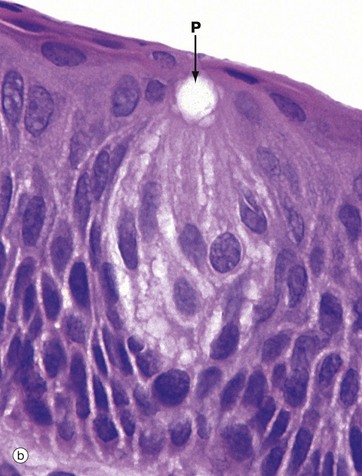
(a) H&E (MP) (b) H&E (HP)
Taste buds, the chemoreceptors for the sense of taste (gustation), are in humans mainly located in the epithelium of the circumvallate papillae of the tongue (see Fig. 13.12), although they are also found scattered in other parts of the tongue, palate, pharynx and epiglottis. In the circumvallate papillae, taste buds face into the deep troughs surrounding the papillae. Serous glands called the glands of von Ebner secrete a serous fluid into the troughs to act as a solvent for taste-provoking substances. The human tongue has approximately 3000 taste buds.
The taste bud is a barrel-shaped organ, occupying the full thickness of the epithelium and opening at the surface via the taste pore P. Each taste bud contains about 50 long, spindle-shaped cells which extend from the basement membrane to the taste pore. Classically, two types of cell are described in the taste bud: light gustatory cells and dark supporting or sustentacular cells. A third cell type, the basal cell, is now generally recognised and may constitute the precursor of one or both of the other cell types. Both gustatory and sustentacular cells have long microvilli extending into the taste pore, which contains a glycoprotein substance, thought to be secreted by the sustentacular cells.
Ultrastructural studies have shown that non-myelinated nerve fibres are associated with both cell types, but there appears to be a more intimate synapse-like relationship between the nerve fibres and the gustatory cells. Although the gustatory cells are thought to be the taste receptors, the sustentacular cells may also serve some receptor function. Like the oral epithelium, all the cells of the taste bud, which represent highly specialised epithelial cells, are renewed continuously, although the gustatory and sustentacular cells are replaced at different rates.
Four taste modalities are recognised: sweet, bitter, acid and salt. Each modality tends to be principally perceived in a specific region of the tongue; however, no structural differences have been demonstrated between taste buds from different areas. The sensations of taste and smell are closely associated, and loss of olfactory sense is accompanied by diminished gustatory perception.
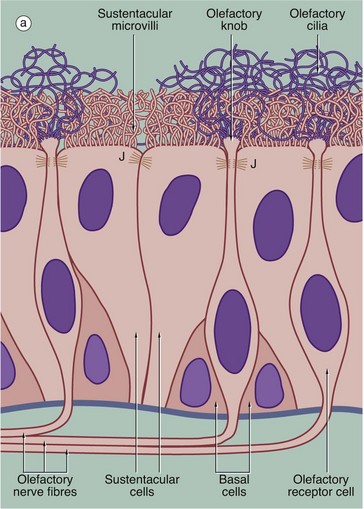
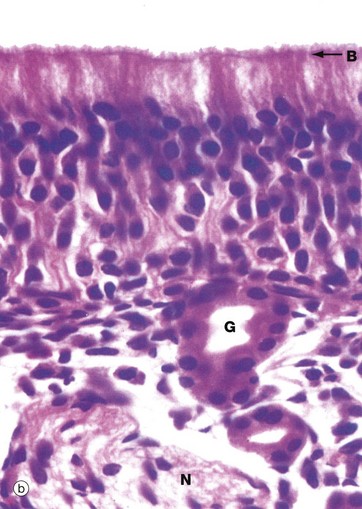
(a) Schematic diagram (b) H&E (HP)
The receptors for the sense of smell are located in a modified form of respiratory epithelium called olfactory epithelium in the nasal cavity; although extensive in some mammals such as the dog, the olfactory epithelium is restricted to a small area in the roof of the nasal cavity in humans. The olfactory epithelium is very tall pseudostratified columnar in form and contains cells of three types: olfactory receptor cells, sustentacular (supporting epithelial) cells and basal epithelial cells.
The olfactory receptor cells are true bipolar neurones (see Fig. 7.2), the cell bodies of which are located in the middle stratum of the olfactory epithelium. A single dendritic process extends from the cell body to the free surface where it terminates as a small swelling, the olfactory knob, which gives rise to about a dozen extremely long modified cilia. These cilia, or olfactory hairs, contain the usual 9 plus 2 arrangement of microtubules in their proximal portion but become thinner distally where they contain variable numbers of single microtubules in different species. The cilia are non-motile and lie flattened against the epithelial surface in the surface mucous layer. The cilia are the sites of interaction between odoriferous substances and the receptor cells. At the basal aspect, each receptor cell gives rise to a single fine non-myelinated axon which penetrates the basement membrane to join the axons of other receptor cells. The bundles of axons pass via about 20 small holes on each side of the cribriform plate of the ethmoid bone to reach the olfactory bulbs of the forebrain where they synapse with second-order sensory neurones.
The supporting or sustentacular cells are elongated with their tapered bases resting on the basement membrane. Many long microvilli extend from their luminal surfaces to form a tangled mat with the cilia of the receptor cells. At the luminal surface, the plasma membranes of the sustentacular and receptor cells are bound together by typical junctional complexes J. The functions of the sustentacular cells are poorly understood, but they probably provide mechanical and physiological support for the receptor cells. The basal cells are small conical cells which appear to be stem cells for both olfactory and sustentacular cells.
In histological section, it is difficult to distinguish individual cell types within the olfactory epithelium; however, the nuclei of sustentacular cells occupy the uppermost stratum, those of the receptor cells the middle stratum and those of the basal cells lie close to the basement membrane. Note the terminal bar B at the luminal surface, representing junctional complexes; note also the fuzzy surface contour representing the tangled meshwork of microvilli and cilia on the surface.
The olfactory epithelium is supported by loose vascular tissue containing bundles of afferent nerve fibres N and numerous serous glands called Bowman’s glands G which produce the watery surface secretions in which odoriferous substances are dissolved.
The Eye
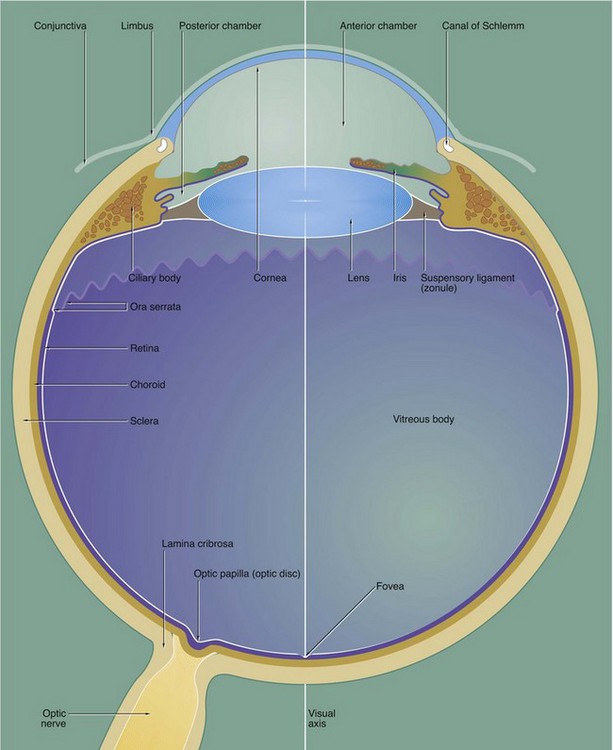
The eye is made up of three basic layers: the outer corneo-scleral layer, the intermediate uveal layer (uveal tract) and the inner retinal layer.
Corneo-scleral layer
The corneo-scleral layer forms a tough fibroelastic capsule which supports the eye. The posterior five-sixths, the sclera, is opaque and provides insertion for the extraocular muscles.
The anterior one-sixth, the cornea, is transparent and has a smaller radius of curvature than the sclera. The cornea is the principal refracting medium of the eye and roughly focuses an image onto the retina; the focusing power of the cornea depends mainly on the radius of curvature of its external surface. The corneo-scleral junction is known as the limbus and is marked internally and externally by a shallow depression. Running from the junction of the cornea and limbus, the surface of the eye is covered by conjunctiva which is reflected into the eyelids.
Uveal layer
The middle layer, the uvea or uveal tract, is a highly vascular layer which is made up of three components: the choroid, the ciliary body and the iris. The choroid lies between the sclera and retina in the posterior five-sixths of the eye. It provides support for the retina and is heavily pigmented, thus absorbing light which has passed through the retina. Anteriorly, the choroid merges with the ciliary body, which is a circumferential thickening of the uvea lying beneath the limbus.
The ciliary body surrounds the coronal equator of the lens and is attached to it by the suspensory ligament or zonule. The lens is a biconvex transparent structure, the shape of which can be varied to provide fine focus of the corneal image upon the retina. The ciliary body contains smooth muscle, the tone of which controls the shape of the lens via the suspensory ligament. The lens, suspensory ligament and ciliary body divide the eye into a large compartment containing a thick gel called the vitreous body and a compartment part in front containing a watery fluid called the aqueous humor.
The iris, the third component of the uvea, forms a diaphragm extending in front of the lens from the ciliary body, so as to incompletely divide the anterior compartment into two chambers; these are known by the terms anterior and posterior chamber. The highly pigmented iris acts as an adjustable diaphragm which regulates the amount of light reaching the retina. The aperture of the iris is called the pupil.
The anterior and posterior chambers contain the aqueous humor, which is secreted into the posterior chamber by the ciliary body and circulated through the pupil to drain into a canal at the angle of the anterior chamber, the canal of Schlemm. The aqueous humor is a source of nutrients for the non-vascular lens and cornea and acts as an optical medium which is non-refractive with respect to the cornea. The pressure of aqueous humor maintains the shape of the cornea.
The large posterior compartment of the eye contains a specialised connective tissue largely composed of a transparent gel known as the vitreous body. The vitreous body supports the lens and retina from within, as well as providing an optical medium which is non-refractive with respect to the lens. In life, the vitreous body contains a canal which extends from the exit of the optic nerve to the posterior surface of the lens; this hyaloid canal represents the course of the hyaloid artery which supplies the vitreous body during embryological development. The vitreous body and hyaloid canal are rarely preserved in histological preparations.
Retinal layer
The photosensitive retina forms the inner lining of most of the posterior compartment of the eye and terminates along a scalloped line, the ora serrata, behind the ciliary body. Anterior to the ora serrata, the retinal layer continues as a non-photosensitive epithelial layer which lines the ciliary body and the posterior surface of the iris.
The visual axis of the eye passes through a depression in the retina called the fovea which is surrounded by a yellow-pigmented zone, the macula lutea. The fovea is the area of greatest visual acuity.
Afferent nerve fibres from the retina converge to form the optic nerve which leaves the eye through a part of the sclera known as the lamina cribrosa. The retina overlying the lamina cribrosa, the optic papilla (optic disc), is devoid of photoreceptors and thus represents a blind spot.
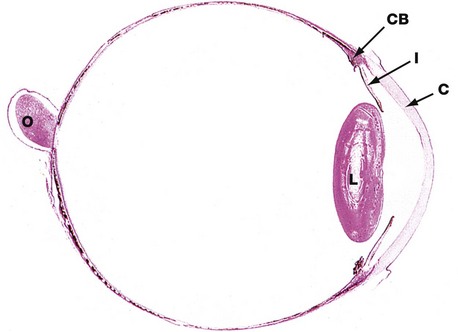
H&E (LP)
This horizontal section shows the relative sizes of the components of the eye. At this magnification, the three layers making up the wall of the globe are not readily distinguishable although, in the wall of the posterior compartment, the middle layer (choroid) is recognisable by its high content of pigment.
The other uveal structures, the ciliary body CB and iris I, are readily visible. The lens L has been artefactually distorted during preparation, and the suspensory ligament by which it is attached to the ciliary body is not preserved. Note the relative thickness of the cornea C.
The optic nerve O is seen to penetrate the sclera medial to the visual axis; the fovea is not present in this plane of section.
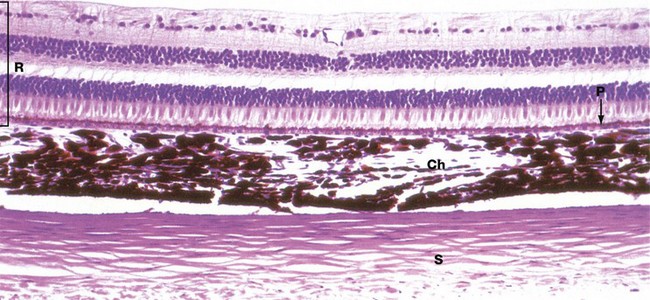
H&E (HP)
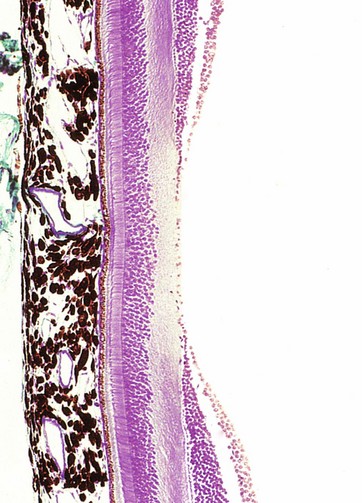
The three layers of the wall of the eye are illustrated in this micrograph.
The inner photosensitive retina is a multilayered structure, the outermost limit of which is defined by a layer of pigmented epithelial cells, the pigment epithelium P.
The choroid Ch is a layer of loose vascular supporting tissue lying between the sclera S externally and the retina R internally. The choroid and retina are separated by a membrane known as Bruch’s membrane which is composed of the basement membranes of the pigmented epithelium of the retina and the endothelium of the choroid capillaries plus intervening layers of collagen and elastin fibres. The blood supply of the uveal layer of the eye is provided by branches of the ophthalmic artery, which penetrates through the sclera. Larger vessels predominate in the superficial aspect of the choroid, with a rich capillary plexus in the deeper aspect providing nourishment for the outer layers of the retina by diffusion across Bruch’s membrane. The choroid contains numerous large, heavily pigmented melanocytes which confer the dense pigmentation characteristic of the choroid. The pigment absorbs light rays passing through the retina and prevents interference due to light reflection.
The sclera consists of dense fibroelastic tissue, the fibres of which are arranged in bundles parallel to the surface. This layer contains little ground substance and few fibroblasts. The sclera varies in thickness, being thickest posteriorly and thinnest at the coronal equator of the globe.
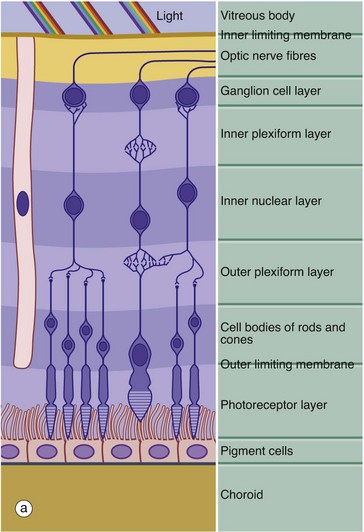
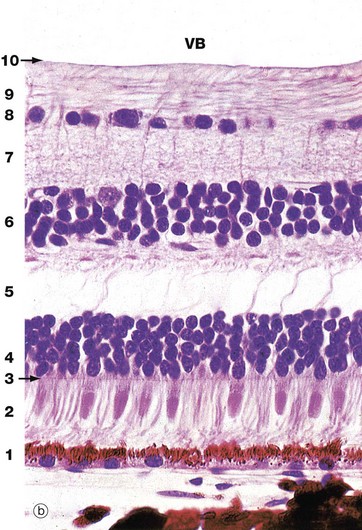
(a) Schematic diagram (b) H&E (HP)
The retina is made up of three cell types: neurones, pigmented epithelial cells and neurone support cells. The neurones are divided into three functional groups: photoreceptor cells (rod cells and cone cells), the cells of afferent fibres passing in the optic nerve and a group of neurones interposed between the first two types which integrate sensory input from the photoreceptors before transmission to the cerebral cortex. The integrating neurones are further subdivided into three types: bipolar cells, horizontal cells and amacrine cells.
Histologically, the retina is traditionally divided into 10 distinct histological layers, as shown in the micrograph; the distribution of the different cell types being illustrated in a highly schematic manner in the diagram.
The outermost layer (1) consists of the pigmented epithelial cells forming a single layer resting on Bruch’s membrane, which separates them from the choroid. The next layer is the photoreceptor layer, made up of the rod and cone processes (2) with a thin eosinophilic structure known as the outer limiting membrane (3) separating them from a layer of densely packed nuclei described as the outer nuclear layer (4). The outer nuclear layer contains the cell bodies of the rod and cone photoreceptors. The almost featureless layer next to this is known as the outer plexiform layer (5) and contains synaptic connections between the short axons of the photoreceptor cells and integrating neurones, the cell bodies of which lie in the inner nuclear layer (6). In the inner plexiform layer (7), the integrating neurones make synaptic connections with dendrites of neurones whose axons form the optic tract. The cell bodies of the optic tract neurones (retinal ganglion cells) comprise the ganglion cell layer (8). Internal to this is the layer of afferent fibres (9) passing towards the optic disc to form the optic nerve. Finally, the inner limiting membrane (10) demarcates the innermost aspect of the retina from the vitreous body VB. Note in the diagram that only bipolar cells are represented in the integrating cell layer; this layer also contains the cell bodies of the horizontal and amacrine cells as illustrated in Fig. 21.8. Note that light impinging on the retina passes through many layers before reaching the photoreceptor cells.
Towards the left of the diagram there is an extremely elongated support cell extending between inner and outer limiting membranes; it has its nucleus in the same layer as the integrating neurones, the inner nuclear layer. These cells, known as Müller cells, are analogous to the neuroglia of the central nervous system and have long cytoplasmic processes which embrace and sometimes even encircle the retinal neurones, filling all the intervening spaces. Müller cells provide structural support and may also mediate the transfer of essential metabolites such as glucose to the retinal neurones.
The outer limiting membrane is not a true membrane but merely represents the line of intercellular junctions between Müller cells and the photoreceptor cells (shown diagrammatically in Fig. 21.7). In contrast, the inner limiting membrane represents the basement membrane of the Müller cells resting on the vitreous body.
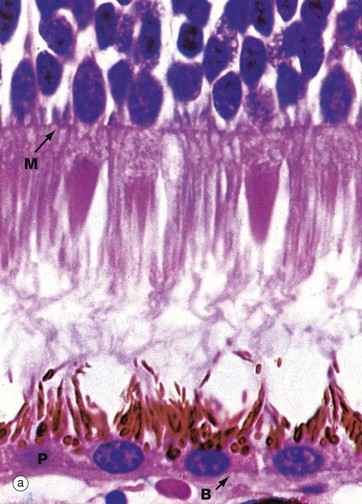
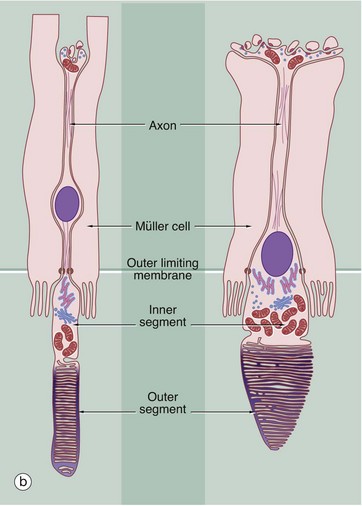
(a) H&E (HP) (b) Schematic diagram
The rod and cone photoreceptor layer of the retina is shown at very high magnification in micrograph (a), the cell bodies of the rod and cone cells lying internal to the outer limiting membrane M. Peripherally, the rods and cones mingle with long microvilli extending from the pigmented epithelial cells P.
As shown in the diagram (b), the rod photoreceptors are long slender bipolar cells, the single dendrite of each cell extending beyond the outer limiting membrane as the rod proper. The rod proper consists of inner and outer segments connected by a thin eccentric strand of cytoplasm containing nine microtubule doublets, similar to those of a cilium but without the inner pair of microtubules. The inner segment contains a prominent Golgi apparatus and many mitochondria. The outer segment has a regular cylindrical shape and contains a stack of flattened membranous discs which incorporate the pigment rhodopsin (visual purple). The membranous discs are continuously shed from the end of each rod and phagocytosed by the pigmented epithelial cells. The discs are continuously replaced from the inner part of the outer segment. In essence, the transduction process involves the interaction of light with rhodopsin molecules which promotes a conformational change in the rhodopsin molecule, thus initiating an action potential. The action potential then passes inwards along the dendrite and axon to the layer of integrating neurones.
Cones are similar in basic structure to the rods, but they differ in several details. The outer segment of the cone is a long conical structure, about two-thirds the length of a rod, and contains a similar number of even more flattened membranous discs. Unlike the situation in the rods, however, the disc membrane is continuous with the plasma membrane so that, on one side, the spaces between the discs are continuous with the extracellular environment. The discs are not shed, although the tips of the cones are invested by processes of pigmented epithelial cells. The cones contain visual pigments similar to rhodopsin, receptive to blue, green and red light, and the mechanism of transduction is probably similar. The bodies of the cone cells are generally continuous with the inner segment of the cone proper, without an intervening dendritic process, and the nuclei of cone cells thus form a row immediately deep to the outer limiting membrane.
As seen in micrograph (a), the pigmented epithelial cells are cuboidal in shape with the nuclei located basally towards Bruch’s membrane B. Apically, the cells are crammed with melanin granules, numerous mitochondria and lipofuscin, a residual product of phagocytosis (see Fig. 1.25). The pigmented cell microvilli, which are 5 to 7 µm long, extend between the photoreceptors and, with electron microscopy, are seen to contain membranous lamellae similar to those in the rod outer segments; these appear to disintegrate as they pass deeper into the pigmented cells. In addition to phagocytosis, the pigmented epithelial cells provide structural and metabolic support for the rods and cones and also absorb light, thus preventing back reflection.
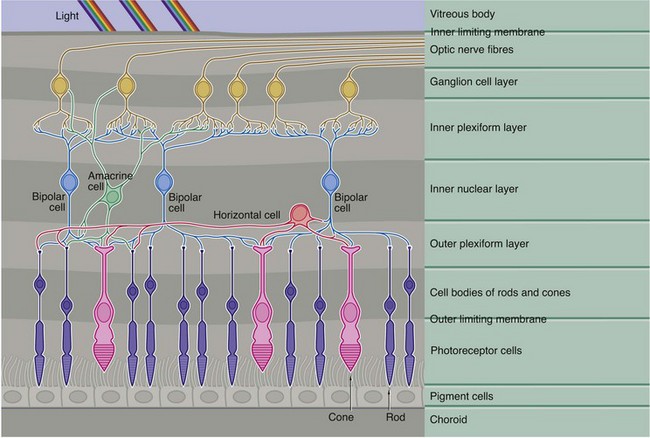
This diagram demonstrates the basic pattern of neuronal interconnections between the photoreceptor cells and the afferent neurones of the optic tract. The interneurones consist of three basic cell types, bipolar cells, horizontal cells and amacrine cells, their cell bodies all being located in the inner nuclear layer (along with those of the supporting Müller cells).
Bipolar cells, the most numerous of the integrating neurones, in general make direct connections between one or more photoreceptors and one or more optic tract neurones, as well as with horizontal and amacrine cells. Horizontal cells have several short processes and one long process, the terminal branches of each making lateral connections between adjacent and more distant rods and cones in the outer plexiform layer. Horizontal cells also synapse with the dendrites of bipolar cells. The amacrine cells have numerous dendrites which make connections with bipolar and optic tract neurones in the inner plexiform layer, as well as making occasional feedback connections with photoreceptors in the outer plexiform layer.
As seen in Fig. 21.7(b) opposite, the axons of the rod photoreceptors terminate in spherical processes into which are invaginated their small number of synaptic connections. In contrast, the cone photoreceptors have a flattened pedicle which accommodates hundreds of intercellular contacts.
In all, there are more than 100 million rods and 6 million cones. The cones are particularly dense in the macula and the immediately surrounding area and, in the fovea itself, the photoreceptors are almost exclusively cones. The density of both rods and cones diminishes towards the retinal periphery. The foveal cones have an almost one-to-one relationship with optic tract neurones, giving maximal visual discrimination. There are only about 1 million optic tract neurones and, the more peripheral the photoreceptors, the greater the number of photoreceptors synapsing with each optic tract neurone. This is consistent with the main function of the more peripheral receptors (predominantly rods), which is for determination of light and dark, rather than fine two-point discrimination.
Masson trichrome (HP)
The fovea is a conical depression in the retina, corresponding to the point where the visual axis of the cornea and lens meets the retina and lying about 4 mm lateral and slightly inferior to the exit of the optic nerve fibres at the optic disc. Consequently, the fovea is the area subject to the least refractory distortion. To complement this, the foveal retina is modified to obtain the maximum photoreceptor sensitivity and is thus the area of the retina with the greatest visual discrimination; however, its function is poor in conditions of low light intensity. Surrounding the fovea is an ovoid yellow area about 1 mm wide called the macula lutea.
As seen in this micrograph, at the fovea the inner layers of the retina are flattened laterally so as to present the least barrier to light reaching the photoreceptors. Retinal blood vessels are absent at the fovea, as can be readily seen with the ophthalmoscope, and the brownish colour of the choroidal melanin shows through the much attenuated retina. At the fovea, the photoreceptors are almost exclusively cones which are elongated and closely packed (approximately 100 000 cones are contained in the fovea). Neuronal interconnections in the bipolar cell layer provide for a one-to-one ratio of these cones to optic nerve fibres, which means that each foveal photoreceptor is individually represented at the visual cortex.![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Basicmedical Key
Fastest Basicmedical Insight Engine
