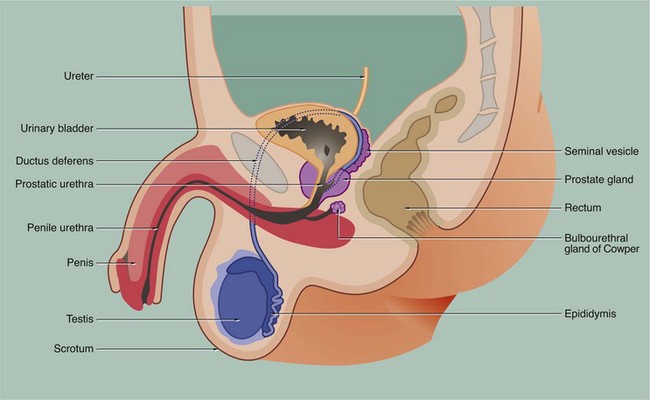
18 • The testes or male gonads, paired organs lying in the scrotal sac, are responsible for production of the male gametes, spermatozoa, and secretion of male sex hormones, principally testosterone. • A system of ducts consisting of ductuli efferentes, epididymis, ductus (vas) deferens and ejaculatory duct collects, stores and carries spermatozoa from each testis. The ejaculatory ducts converge on the urethra, from which spermatozoa are expelled into the female reproductive tract during copulation. • Two exocrine glands, the paired seminal vesicles and the single prostate gland, secrete a nutritive and lubricating fluid medium called seminal fluid in which spermatozoa are conveyed to the female reproductive tract. Semen, the fluid expelled during ejaculation, consists of seminal fluid and spermatozoa, plus some desquamated duct-lining cells. • The penis is the organ of copulation. A pair of small accessory glands, the bulbourethral glands of Cowper, secrete a fluid which lubricates the urethra for the passage of semen during ejaculation. FIG. 18.2 Testis FIG. 18.3 Testis, monkey In all somatic cells, cell division (mitosis) results in the formation of two daughter cells, each one genetically identical to the mother cell. Somatic cells contain a full complement of chromosomes (the diploid number) which function as homologous pairs (see Ch. 2). The process of sexual reproduction involves the fusion of specialised male and female cells called gametes to form a zygote, which has the diploid number of chromosomes. Each gamete contains only half the diploid number of chromosomes, one representative of each pair; this half complement of chromosomes is known as the haploid number. The production of haploid cells involves a unique form of cell division called meiosis, which occurs only in the germ cells of the gonads during the formation of gametes; meiotic cell division is thus also called gametogenesis. Meiosis involves two cell division cycles, of which only the first is preceded by duplication of chromosomes (see Ch. 2). Thus, meiotic cell division of a single diploid germ cell gives rise to four haploid gametes. In the male, each of the four gametes undergoes morphological development into a mature spermatozoon. In contrast, in the female, unequal distribution of the cytoplasm during meiosis results in one gamete gaining almost all the cytoplasm from the mother cell, while the other three acquire almost no cytoplasm; the large gamete matures to form an ovum and the other three, called polar bodies, degenerate. The primitive germ cells of the male, the spermatogonia, are present only in small numbers in the male gonads before sexual maturity. After puberty, spermatogonia multiply continuously by mitosis to provide a supply of cells which then undergo meiosis to form male gametes. In contrast, the germ cells of the female, called oogonia, multiply by mitosis only during early fetal development, thereby producing a fixed complement of cells with the potential to undergo gametogenesis. Gametogenesis in the female is discussed more fully in Ch. 19. The production of male gametes is called spermatogenesis and the subsequent development of the male gamete into a motile spermatozoon is called spermiogenesis, the whole process taking approximately 70 days; both these processes occur within the testes, although final maturation of spermatozoa occurs in the epididymis. FIG. 18.4 Seminiferous tubules In this micrograph of normal testis at medium power, note the seminiferous tubules Se cut in various planes of section, giving round and ovoid profiles. Between the seminiferous tubules the interstitium I contains Leydig cells (which cannot be discerned at this magnification) and small capillaries C. Larger arteries and veins are found in the fibrous septa that divide the organ into lobules. FIG. 18.5 Seminiferous tubule FIG. 18.6 Spermiogenesis 3. Meanwhile, both centrioles migrate to the end of the cell opposite to the acrosomal head cap; the centriole aligned parallel to the long axis of the nucleus elongates to form a flagellum which has a basic structure similar to that of the cilium (see Fig. 5.13). The mitochondria become arranged in a helical manner around the fibrils which surround the first part of the flagellum.
Male reproductive system
Introduction
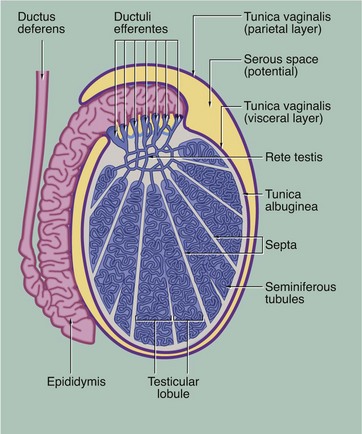
During embryological development, each testis with the first part of its duct system, blood vessels, lymphatics and nerves descends from the posterior wall of the peritoneal cavity to the scrotum. During migration, the testis carries with it an investing layer of peritoneum so that in the scrotum the testis is almost completely surrounded by a double layer of mesothelium, enclosing a potential space. This double lining is called the tunica vaginalis and, like the pleura, consists of visceral and parietal layers, separated by a thin layer of serous fluid. The fluid is secreted by the mesothelial cells and acts as a lubricant, allowing the testis to move freely in the scrotal sac. The visceral layer of the tunica vaginalis rests on the capsule of the testis, the tunica albuginea, which gives rise to numerous incomplete collagenous septa. These divide the testis into about 250 testicular lobules. Within each lobule, there are one to four highly convoluted tubes, the seminiferous tubules, in which spermatozoa are produced. The seminiferous tubules converge upon a plexus of channels, the rete testis. From the rete testis, 15 to 20 small ducts called the ductuli efferentes carry spermatozoa to the extremely tortuous first part of the ductus deferens, which is known as the epididymis.
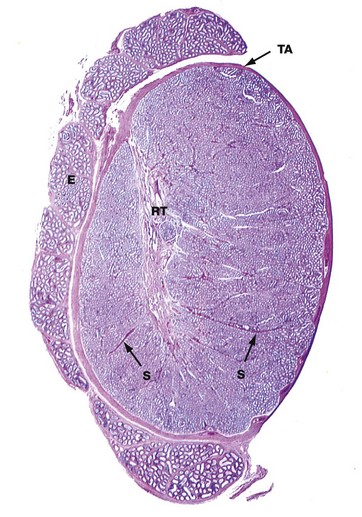
H&E (LP)
This micrograph illustrates the macroscopic features of a testis; cut in the sagittal plane, it shows the relationship to the epididymis E, which lies on its posterior aspect. The testis is packed with coiled seminiferous tubules which can just be seen in various planes of section at this magnification. Groups of up to four seminiferous tubules are segregated into testicular lobules by fine interlobular septa S.
The dense fibrous capsule which invests the testis, and which is continuous with many of the interlobular septa, is called the tunica albuginea TA. It contains fibroblasts and abundant myofibroblasts and smooth muscle cells, particularly in the posterior aspect close to the rete testis, which subject the seminiferous tissue to rhythmic contractions. Scattered Leydig cells are also found within the tunica albuginea. The deepest layer of the tunica albuginea consists of loose connective tissue containing blood and lymphatic vessels, sometimes called the tunica vasculosa.
Spermatozoa pass from the seminiferous tubules into the rete testis RT, which is connected to the epididymis via the ductuli efferentes at the upper posterior pole of the testis; the ductuli are not included in the plane of this section. The epididymis is a tightly coiled tube which forms a compact mass extending down the whole length of the posterior surface of the testis and is the major site of storage of newly formed spermatozoa. At the lower pole of the testis, the epididymal tube becomes continuous with the relatively straight ductus (vas) deferens, not seen in this section.
Gametogenesis
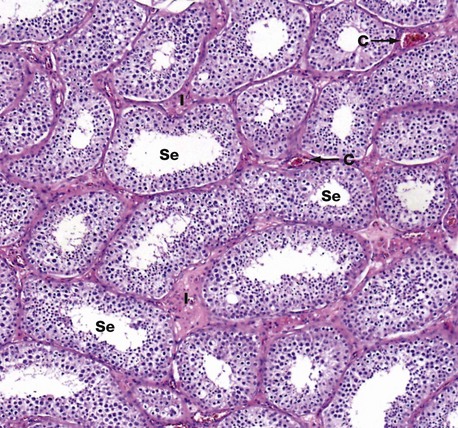
H&E (MP)
This micrograph illustrates seminiferous tubules cut in various planes of section. The seminiferous tubules are highly convoluted and are lined by:
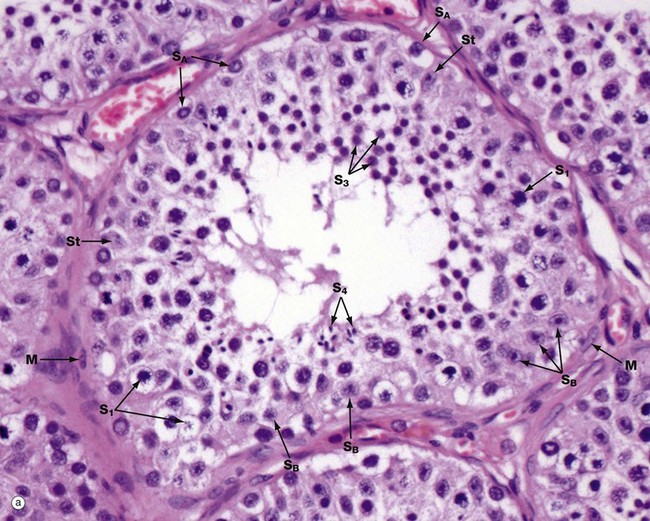
(a) H&E (HP) (b) Diagram
Micrograph (a) illustrates an adult seminiferous tubule cut in transverse section. The processes of spermatogenesis and spermiogenesis are synchronised, with waves of activity occurring sequentially along the length of each tubule. Thus in a single cross-section of a tubule, not all development phases will be represented (b).
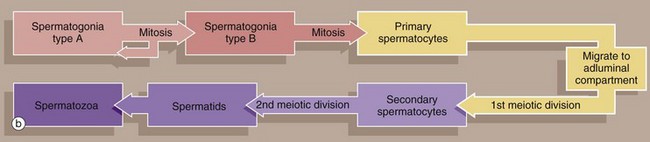
The undifferentiated diploid germ cells, found in the basal compartment of the seminiferous tubule, are called type A spermatogonia. These go through several cycles of mitosis to produce further type A spermatogonia, which maintain the germ cell pool, and type B spermatogonia, which are committed to production of spermatozoa. Spermatogonia type A SA are characterised by a large round or oval nucleus with condensed chromatin; peripheral nucleoli and a nuclear vacuole may be prominent. Spermatogonia type B SB have dispersed chromatin, central nucleoli, and no nuclear vacuole. Both types of spermatogonia have sparse poorly stained cytoplasm.
Type B spermatogonia undergo further mitotic divisions to produce primary spermatocytes. These migrate to the adluminal compartment of the seminiferous tubule before commencing the first meiotic division. Primary spermatocytes S1 are readily recognised by their copious cytoplasm and large nuclei containing coarse clumps or thin threads of chromatin; dividing cells may be seen. In humans, the first meiotic division cycle takes approximately 3 weeks to complete, after which time the daughter cells become known as secondary spermatocytes. The smaller secondary spermatocytes rapidly undergo the second meiotic division and are therefore seldom seen.
The gametes thus produced, called spermatids S3, then proceed through the long maturation process known as spermiogenesis to become recognisable as spermatozoa. During this process, the nuclei of the spermatids assume the small pointed form of spermatozoa S4 (see Fig. 18.7). Examination of different sections of the tubules of a normal testis shows about half the spermatogenic cells to be in the late spermatid stage.
During the developmental process, the cells of the spermatogenic series are supported by Sertoli cells St, whose nuclei are usually found towards the basement membrane of the seminiferous tubule. The Sertoli cell nucleus is typically triangular or ovoid in shape with a prominent nucleolus and dispersed chromatin.
The basal layer of germinal cells is supported by a basement membrane which is surrounded by a lamina propria containing several layers of spindle-shaped myofibroblasts M and fibroblasts.
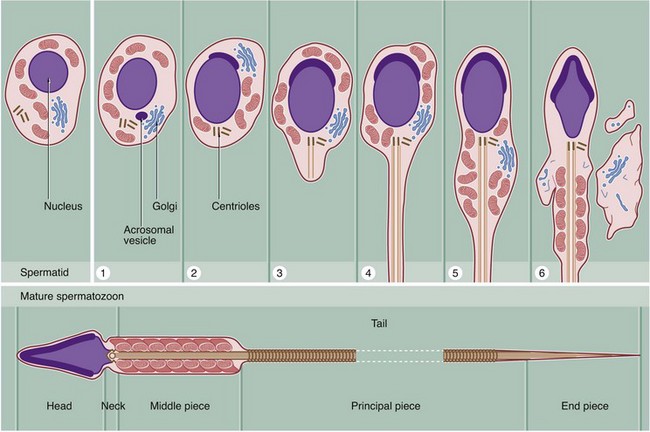
Spermiogenesis is the process by which spermatids, the gametes produced by meiotic division, are transformed into motile mature spermatozoa. This involves the following major stages:
The structure of fully formed spermatozoa varies in detail from species to species but conforms to the basic structure seen in this diagram of a human spermatozoon.
Throughout the entire developmental process from spermatogonia to spermatozoa, hundreds of spermatids remain connected to one another by narrow cytoplasmic bridges which only break down upon release of spermatozoa into the lumen of the seminiferous tubule. This explains the synchronous development of spermatozoa at any one part of the tubule.
Sertoli cells are important in the regulation of spermatogenesis and spermiogenesis. Sertoli cells form tight junctions with each other as well as with the developing germ cells. It is well established that high concentrations of androgen hormones, secreted by Leydig cells of the testicular interstitium (see Fig. 18.10), are essential for production and maturation of spermatogenic cells. Sertoli cells secrete an androgen-binding protein which transports testosterone and dihydrotestosterone to the lumen of the seminiferous tubule. These hormones are also necessary for function of the epithelium of the rete testis and epididymis; production of this binding protein is believed to be dependent on the pituitary gonadotrophin follicle stimulating hormone (FSH).![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Basicmedical Key
Fastest Basicmedical Insight Engine